第三节 痴呆症状有关的神经生理学基础
大脑不断地接受从机体各部位感受器传来的信息,信息到达大脑组织后,加工形成感知觉,经整合后进而制订行动计划,组合各种有效的行为活动。大脑之所以有如此复杂的功能是因为人类大脑内有数以千亿计的神经元,它们相互连接构成了一个极其复杂的网络系统,这个由神经元组成的特殊信号转导网络系统调节着相应的行为,因此,人类行为的复杂性主要决定于大量神经元精确排列的神经环路。
神经调节的基本活动方式是反射,反射是由神经元按照一定连接调制的感觉—运动环路实现的(图4-3-1)。人类的大脑高度发展,除感觉与运动外,还能完成更复杂的高级功能活动,如学习和记忆、思维和判断、语言和其他认知活动等。大脑皮质是反射的最高级中枢,皮质的Ⅰ~Ⅳ层主要接受传入冲动。由丘脑上行的特异传入纤维,主要进入第Ⅳ层与星形细胞形成突触,星形细胞的轴突又与其他皮质细胞建立广泛的突触联系,从而对皮质的各种信息进行分析整合,作出相应的反应;由大脑半球同侧或对侧联合来的传入纤维则进入皮质第Ⅱ、Ⅲ层,与锥体细胞形成突触。大脑皮质的传出纤维分投射纤维和联合纤维两种,投射纤维由第Ⅴ层的锥体细胞和大梭形细胞发起,下行至脑干及脊髓;联合纤维起自第Ⅲ、Ⅳ、Ⅴ的锥体细胞和梭形细胞,分布于皮质的同侧或对侧脑区。皮质的第Ⅱ、Ⅲ、Ⅳ层细胞主要与各层互相联系,构成复杂的神经微环路,对各种信息进行分析、整合和储存。由于大脑皮层联系的复杂性和广泛性,使皮层具有分析和综合的能力,从而构成了人类思维活动的物质基础。
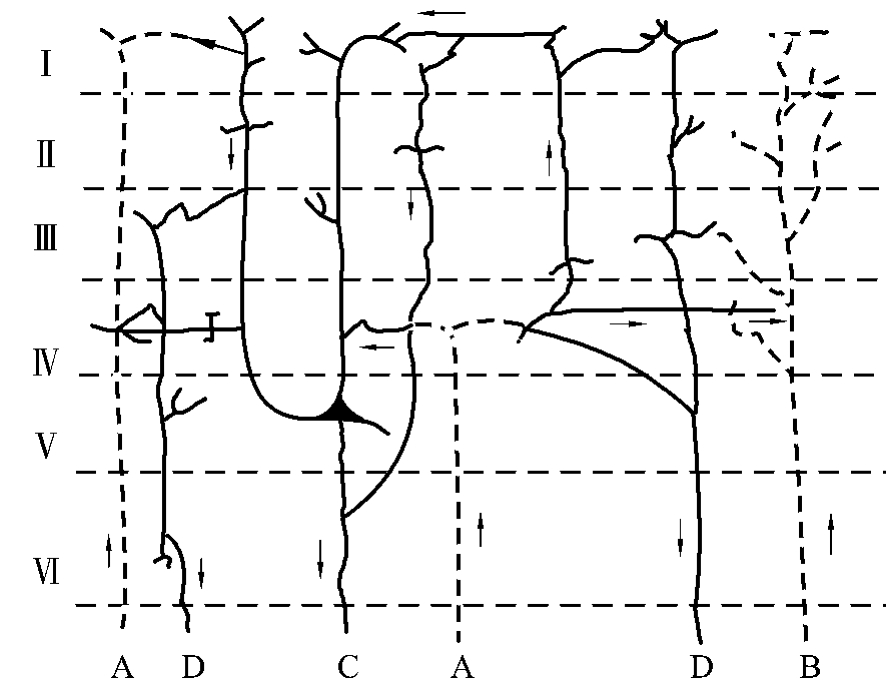
图4-3-1 大脑皮层内微环路简图。A.特异传入纤维; B.联合传入纤维;C.投射传出纤维;D.联合传出纤维
一、学习和记忆的生理学基础与记忆障碍
(一)学习和记忆
学习是指人或动物依赖于经验来改变自身行为以适应环境的神经活动过程。记忆是将学习到的信息进行储存和“读出”的神经活动过程。因此学习和记忆是相互关联密不可分的两个活动过程,可将其分为获得(acquisition)、巩固(consolidation)和再现(retrieval)三个阶段。获得是学习阶段,是动物或人感知外界事物或接受外界信息后,通过感觉传入系统向中枢传递信号的过程。在这一阶段,传入的信号能被大脑获得、并进一步加工处理,仅占机体能感受到的大量信息中的很少部分。巩固阶段,包括信息在脑内识别、复制、编码、储存等过程。信息在脑内保存时间的长短,与传入的信息对个体的意义及应用有关,意义重大或反复应用的信息可较长时间地储存,而那些不能巩固的信息就会丢失,出现遗忘。可见,巩固是学习过程中的讯号转入记忆系统的过程。再现阶段是把已在脑内编码储存的信息通过回忆或相应刺激再次浮现并读出的过程。根据所获得的信息不同,再现可包括场景画面再现、事件经过再现、语言和声音再现及有规律行为活动的再现等。
1.学习的形式
(1)非联合型学习 非联合型学习也称简单学习,是指在学习过程中引起反应的刺激是单一的,刺激与反应之间不存在某种明确的联系或联合。它具有习惯化和敏感化特征,其机制与习惯化和敏感化等可塑性改变有关。
当非伤害性刺激重复作用于机体时,其引起的反射性效应会逐渐减弱,这个过程称为习惯化。如刚到一个新的环境,听到钟表发出的“嘀嗒”声,可能使人难以入睡,但几天过后,该钟表的声音便不再被注意到。通过习惯化使人或动物学会辨别某种刺激的能力,在以后的生活中忽略这些对我们生存没有明确意义的信息,这有利于机体接受其他各种重要的信息。
当人或动物受到某种强烈的或伤害性刺激后,机体对其他刺激的反应性增强的过程称为敏感化。例如:当动物受到强烈的痛觉刺激后,它对温和的触觉刺激也会产生强烈的反应。敏感化的形成不依赖于强刺激和弱刺激之间的配对,在时间上也不需要二者的结合,故又称为假性条件化。敏感化的意义与习惯化相反,它使个体学会了对某些强烈的或伤害性刺激的注意,有利于躲避该刺激。
(2)联合型学习 “联合”是指在学习过程中需要两种不同的刺激按照一定次序进行配对,由此在脑内逐渐形成相互联系。在联合型学习中,人或动物学会了强刺激和弱刺激之间或行为与某种刺激间的联系。联合学习有多种形式,其中最著名的是经典条件反射和操作式条件反射。
1)经典条件反射的建立:由条件刺激引起的反射,称为条件反射,又称巴甫洛夫条件反射。经典巴甫洛夫实验中,狗学会了听到铃声预示着即将进食。它是由条件刺激(如铃声)和非条件刺激(如食物)在时间上的多次结合而建立起来的,这个过程称为强化。只有通过强化,才能建立条件反射。条件反射是对提前到来信号(条件刺激)建立的反射。所以,经典的条件反射大大提高了机体的预见性,从而增强了机体的适应能力。
若多次只给予条件刺激(如铃声),不给予非条件刺激(如进食)强化,已形成的条件反射会逐渐减弱,最后完全消失,这称为条件反射的消退。消退并不是遗忘,它是一个新的学习过程,因此所有动物和人都有建立经典条件反射的能力,可见该能力对动物的生存极其重要。
2)操作式条件反射:该反射活动使人和动物建立了刺激与行为间的联系。它要求动物或人通过学习完成一定的操作。一个典型的实例是将一个饥饿的大鼠放入实验箱内,大鼠必须通过自己完成某种运动或操作(如压杠杆)后才能得到非条件刺激(如食物)的强化。大鼠从而学会了在它许多行为中,有一种是获得食物的压杠杆行为,此后大鼠一旦饥饿就去压杠杆,这种学习过程使动物学会了用自己的一种行为预示着强化刺激的到来。操作性条件反射常用于动机行为的研究。在形成简单的操作性条件反射后,还可进一步联合其他的刺激,以形成更复杂的操作性条件反射。动物得到食物是一种奖赏性刺激,称为“趋向性”条件反射。在训练动物时得到惩罚(例如电击)而不去踩杠杆,形成抑制性条件反射,称为“回避性”条件反射。
2.人类的条件反射和两种信号系统学说条件反射是对信号发生反应的神经过程。大脑皮层对不同信号发生反应的功能系统分为两种信号系统。把现实具体的信号称为第一信号,大脑皮层对第一信号发生反应的功能系统称为第一信号系统,人类和动物均以第一信号作为条件刺激发生条件反射,如饥饿时看到或嗅到食物即可引起唾液分泌。
具体信号的抽象概括为第二信号,大脑皮层对第二信号发生反应的功能系统称为第二信号系统,只有人类才具有语言和文字功能,第二信号系统是人类区别于动物的主要特征。第二信号的所谓“抽象”,是指语词所包含的内容,而不是语言中声波的物理振动和文字中笔画的空间排列。因此人类借助于语词表达思想,对一切现实事物和现象进行抽象概括,形成概念并进行推理,不断扩大认识能力,从而更深刻地认识和改造客观世界。
3.记忆的形式 记忆可根据信息的类型、储存和读出的方式以及保留时间的长短来进行分类。
(1)根据记忆的储存和回忆方式分为陈述性记忆(情节记忆和语义记忆)、非陈述性记忆两类。
1)陈述性记忆:情节记忆和语义记忆,统称为陈述性记忆(declarative memory),都依赖于海马—间脑内侧系统的调控。
陈述性记忆是对本人所经历和学习的事件和情景进行编码、贮存并回忆、再现的过程。记忆内容常常较生动,如人生中经历的重大事件或参加一次有特殊意义活动的场景,在很多年以后仍能回忆和再现,并用语言表达出来,属于陈述性记忆中的情节记忆(episodic memory)。对周围世界一般知识如文字、语言和数学定义等的陈述性记忆,称为语义记忆(semanticmemory),与获得记忆时的情景无关。
2)非陈述性记忆:工作记忆属非陈述性记忆(nondeclarative memory)。心理学家将有关技能的记忆称为非陈述性记忆,是对一系列规律性操作程序的记忆,又称反射性记忆(reflexive memory)。主要表现是行为的改变,如弹钢琴、游泳、体操运动员优美而连贯的动作技巧的学习,以及对程序和规则学习的记忆,称为程序性记忆(proceduralmemory)。
4.人类的记忆过程 按照记忆的发展过程,又可将记忆分成四个阶段,即感觉性记忆、第一级记忆、第二级记忆和第三级记忆。前两个阶段相当于短时程记忆,后两个阶段相当于长时程记忆(图4-3-2)。
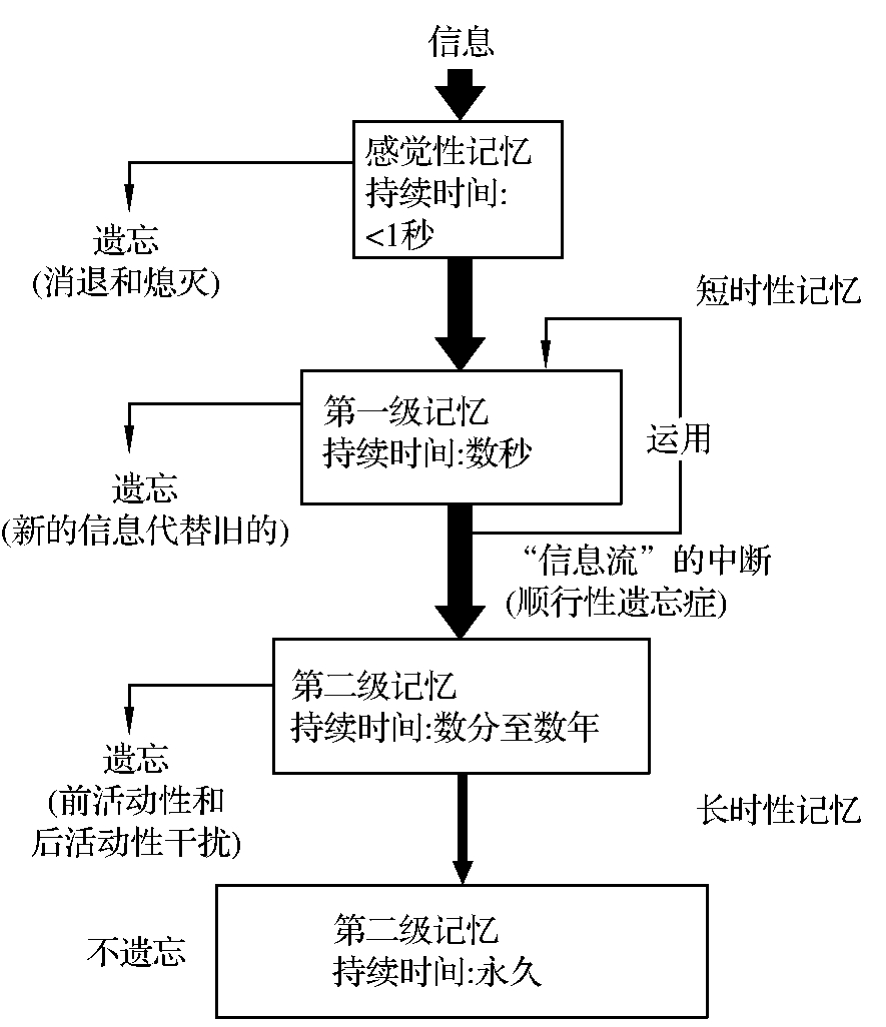
图4-3-2 记忆过程的四个连续阶段
感觉性记忆属于短时程记忆,指的是对短时间内(几秒钟到几分钟)输入信息的记忆能力,是机体经感觉系统获得的信息在脑的感觉代表区储存的阶段。所有进入机体的信息都要经过这一阶段。信息在此阶段保留的时间很短,一般不超过1s,信息的储存是不牢固的。如果对这些短暂的感觉性的信息加以注意和加工处理,感觉性信息将转换成表达性符号(如临时打一个电话时,将查出的号码念一遍),感觉性记忆便可转入第一级记忆。
第一级记忆属于短时程记忆,即时应用性信息可进入此过程,信息在此阶段保留的时间仍很短,从数秒到数分钟,其信息记忆容量也有限。例如临时拨打一个陌生的电话号码,当时可以记住它,打完电话几分钟后,该电话号码就会忘掉。只有受到注意和经过复习的小部分信息才能保持在短时程记忆中。短时程记忆有多种表现形式,如影像记忆、即刻记忆和工作记忆或操作记忆。
第二级记忆属于长时程记忆。如果某些信息需要反复应用,信息便在第一级记忆中循环,并可转入第二级记忆中。信息在此可以保存数分至数年,因此也是一个贮量较大的系统。人体需要保存的信息(反复应用性信息),大部分都储存在此记忆之中。
第三级记忆也属于长时程记忆,它是在第一记忆的基础上,通过反复运用、强化而转入第二记忆。在第二记忆中,信息可因前面的信息或后来的信息干扰而造成遗忘,分别称为前活动性干扰和后活动性干扰。在克服了干扰后,则转入第三记忆,成为永久性记忆。
长时程记忆是一个庞大而持久的储存系统,随着新的记忆不断储存,有些旧的记忆也在不断丢失。在个体生存过程中,只有那些重大事件或终年累月运用的信息,如自己的名字和常年进行的操作手艺等,才能达到永久记忆而终身不忘。
5.学习和记忆的机制
(1)学习与记忆的脑功能定位 PET和fMRI发现程序记忆储存在纹状体、运动皮层、小脑学习和记忆过程在脑区有一定的功能定位。与学习和记忆有密切关系的脑区有大脑皮层联络区、海马及其邻近结构、杏仁核、丘脑及脑干网状结构等部位(图4-3-3)。顶叶皮层可能储存有关地点的影像记忆。额叶皮层在短时程记忆中发挥重要作用。
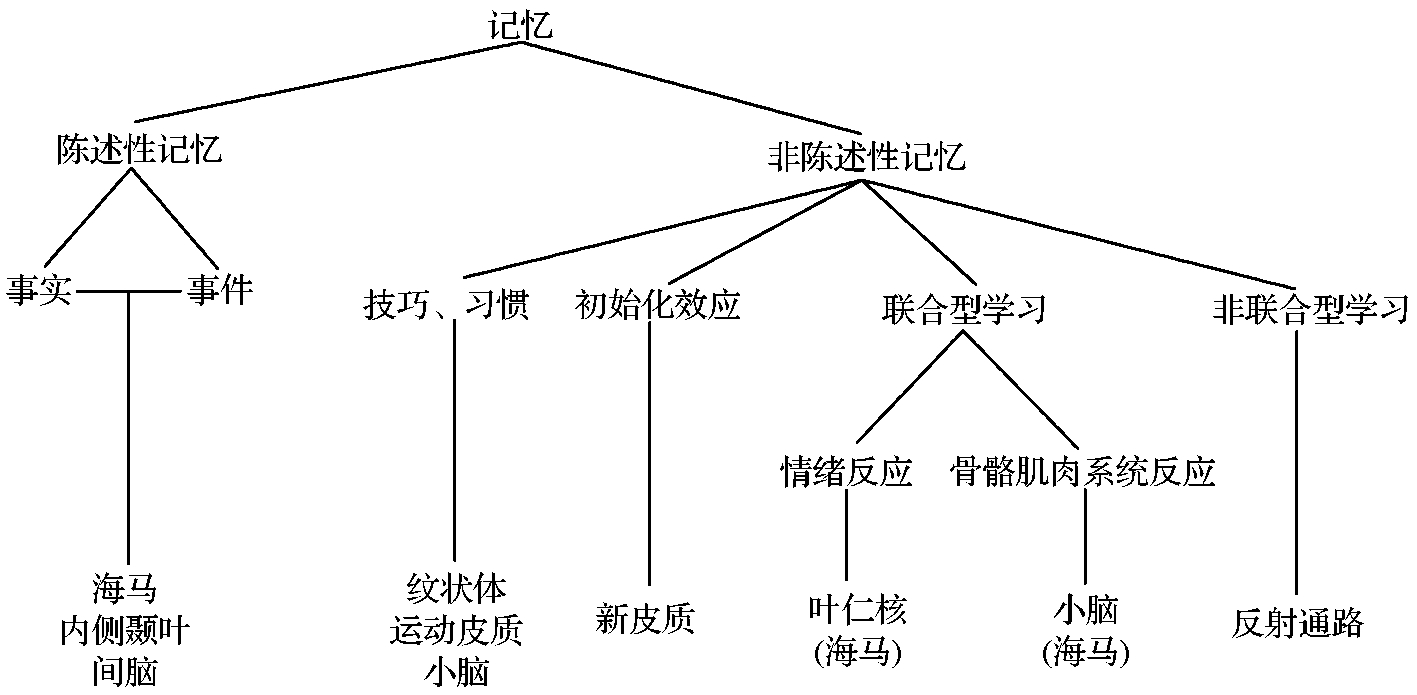
图4-3-3 记忆的分类及与其相关的脑区
运动学习是一种典型的程序性记忆,其形成过程主要与大脑皮层、边缘系统经伏隔核到纹状体苍白球的通路有关(图4-3-4)。
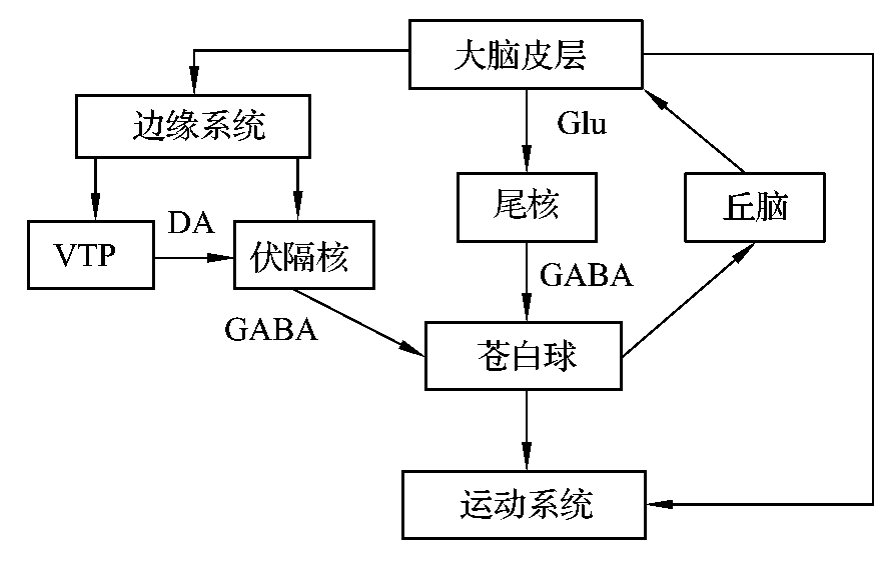
图4-3-4 非陈述性记忆神经调控回路的示意图注:Glu:谷氨酸DA:多巴胺GABA:γ-氨基丁酸
中、短时程的陈述性记忆需要大脑皮层联络区及海马回路的参与。视、听、触、压觉冲动传入大脑皮层后到达皮层联络区,味、嗅觉冲动经颞叶和额叶的边缘皮层到达皮层联络区,继而两路信息再经内侧颞叶边缘系统、丘脑内侧核团、额叶腹内侧部进入基底—前脑胆碱能系统,最后回到大脑皮层联络区(图4-3-5)。
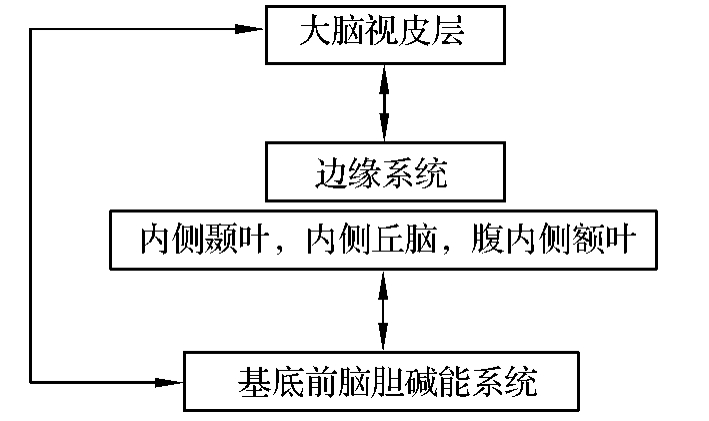
图4-3-5 陈述性记忆神经回路的示意图
在陈述性记忆分子机制的临床观察和动物实验研究中发现,海马参与了长时陈述记忆的形成。电生理实验发现海马神经元间的突触联系具有可塑性。海马的早期和晚期长时程增强可能是陈述性记忆的突触机制。1973年Bliss及Lomo首先在麻醉兔的脑海马穿通纤维上给予一组高频电刺激,记录齿状回颗粒细胞放电,发现单个刺激引起的群体锋电位(population spike,PS)和群体兴奋性突触后电位(population EPSP)的振幅增大,群体锋电位的潜伏期缩短。这种易化现象持续的时间可长达10 h以上。Bliss将这种单突触诱发反应的长时程易化称为海马神经元的长时程增强(LTP)。1983年,Colingridge等人发现NMDA受体通道复合体在LTP过程中起重要作用,进一步深化了对LTP在大脑学习记忆中作用的理解。
海马的LTP具有三个基本特征:①协同性,诱导LTP需要很多纤维同步放电;②联合性,在海马的任何神经纤维通路上LTP的形成需要一组高频刺激诱发突触后神经元(CA1)和突触前神经元(CA3)的同步放电;③特异性,所诱导的LTP对诱发过程中的被激活的输入路径是特异的,在其他输入路径上不产生LTP。
海马有三条主要兴奋性通路:①前穿通纤维到齿状回颗粒细胞;②从颗粒细胞到CA3区锥体细胞的苔状纤维;③从CA3到CA1区锥体细胞Schaffer侧支(图4-3-6)。在这三个通路的单突触连接的任何神经纤维上都可引导出LTP,但不同部位的LTP可能有不同的神经机制:例如CA1区的LTP主要通过突触后谷氨酸(NMDA)受体,CA3区的Mossy纤维上LTP发生于突触前,不依赖于NMDA受体。
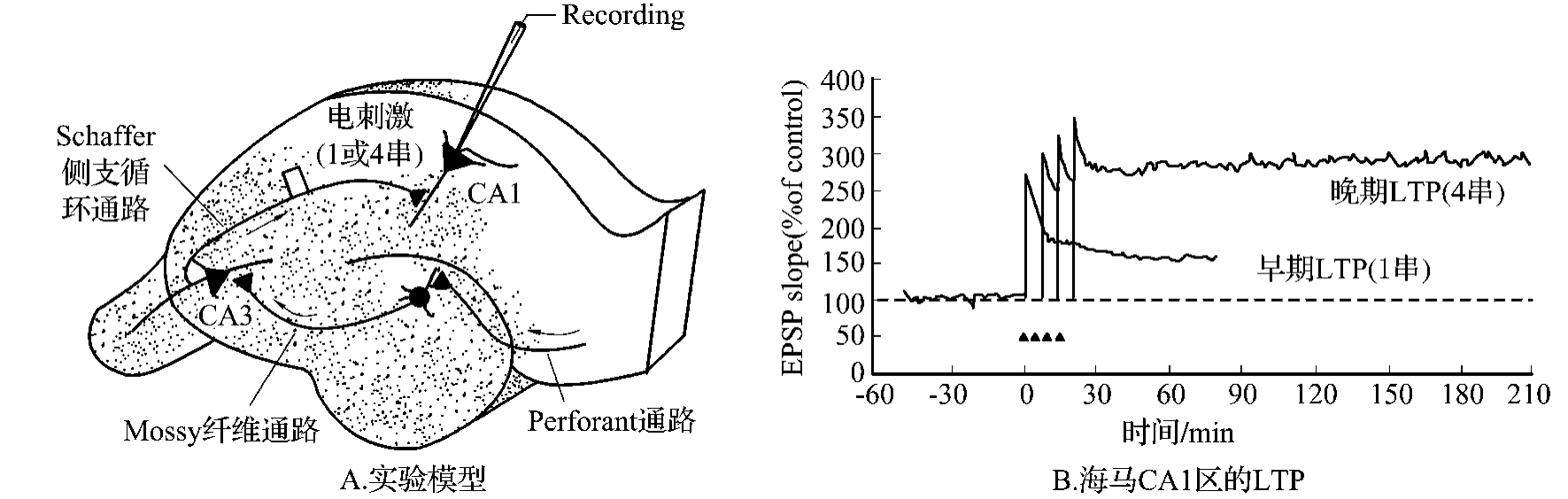
图4-3-6 A.电刺激海马Schaffer侧支纤维,记录CA1区锥体细胞的电活动;B.海马早期和晚期LTP的比较
谷氨酸受体可分为促离子型受体和促代谢型受体两种类型。促离子型受体可再分为海人藻酸(kainic acid或kainate,KA)受体、AMPA(α-amino-3-hydroxy-5-methyl-4-isoxazoleproprionate)受体和NMDA(N-methyl-D-aspartate)受体三种类型。KA受体和AMPA受体过去合称为非NMDA受体。
长时程增强(LTP)可见于神经系统的许多部位,但研究最多最深入的是海马。人们常根据NMDA谷氨酸受体阻断剂的作用将LTP分为两类:NMDA受体依赖的LTP;非NMDA受体依赖的LTP。目前对NMDA受体依赖的LTP的机理研究的较多。
在正常低频突触传递时,突触前膜释放的谷氨酸可同时作用于突触后膜上NMDA和非NMDA受体,但只能激活非NMDA受体出现较弱的去极化反应;因非NMDA受体对谷氨酸的反应较快,其耦联通道的电阻较低(尤其是KA受体),激活时主要增加对Na+和K+的通透性,产生突触后电位。NMDA受体对谷氨酸的反应较慢,其耦联通道的电阻较高,激活时主要增加对Na+、K+和Ca2+的通透。
NMDA受体是一种独特的双重门控通道,其特点有:①既受谷氨酸递质门控也受电压门控调节;②膜外侧存在能与甘氨酸结合的位点,甘氨酸与之结合不仅为谷氨酸产生兴奋效应所必需,而且能增其耦联通道的开放频率;③通道内某一点可与Mg2+结合而将通道阻塞,Mg2+阻塞通道是电压依赖型的,细胞超极化程度越高,Mg2+进入NMDA受体耦联通道越多,阻滞Ca2+-Na+-K+通道程度越高。当CA1细胞与CA3细胞同步放电时,膜去极化达到一定程度后,才能移开堵塞通道的Mg2+,使结合了Glu的NMDA通道打开(图4-3-7);④通道内某些受点可与苯环立啶和氯胺酮等药物结合而发生变构,从而降低对Ca2+-Na+-K+通透性;⑤NMDA受体以海马和大脑皮层分布最多,绞状体和隔较多。
一组高频刺激可使突触后膜的去极化达到一定程度,使Mg2+对NMDA通道的阻塞效应解除,从而打开NMDA通道,出现Ca2+内流,Ca2+进入CA1细胞内与钙调蛋白(CaM)结合,进一步激活Ca2+-CaM依赖性激酶、PKC和酪氨酸激酶产生逆行信使(NO、CO等),增加突触前神经递质的释放和突触后受体的敏感性,导致持续1~3 h的早期LTP(early LTP,E-LTP)产生,相当于短时记忆。
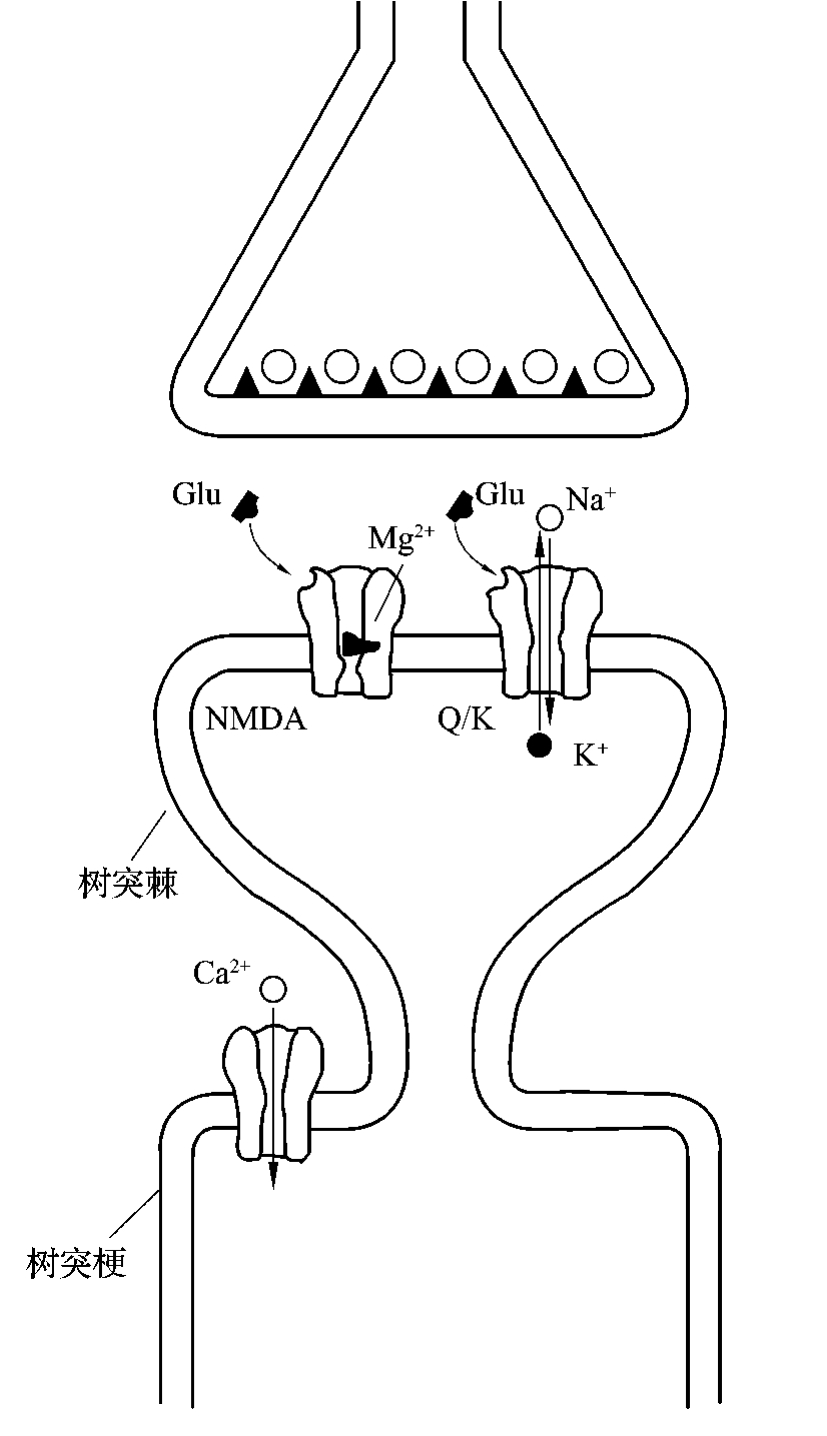
图4-3-7 NMDA受体双重门控通道模式图
脑缺血时,兴奋性氨基酸—谷氨酸含量急剧升高,过量的谷氨酸通过激活N-甲基-D-天冬氨酸(NMDA)受体,使Ca2+通道开放,胞外钙离子大量内流,可激活细胞膜上的磷脂酶A等酶系,使体内自由基的生成和清除失衡,导致脂质过氧化损伤,从而引起神经元迟发性损伤。
给予多组高频电刺激,则可以引发持续24 h以上的晚期LTP(late LTP,L-LTP),相当于长时程记忆,与长时非陈述记忆相似(图4-3-8)。晚期LTP依赖于mRNA和新蛋白质的合成,并需要激活cAMP-PKA-丝裂原激活的蛋白激酶(mitogen-activated protein kinase,MAPK)-cAMP反应元件结合蛋白(cAMP response element binding protein,CREB-1)信息传导途径。
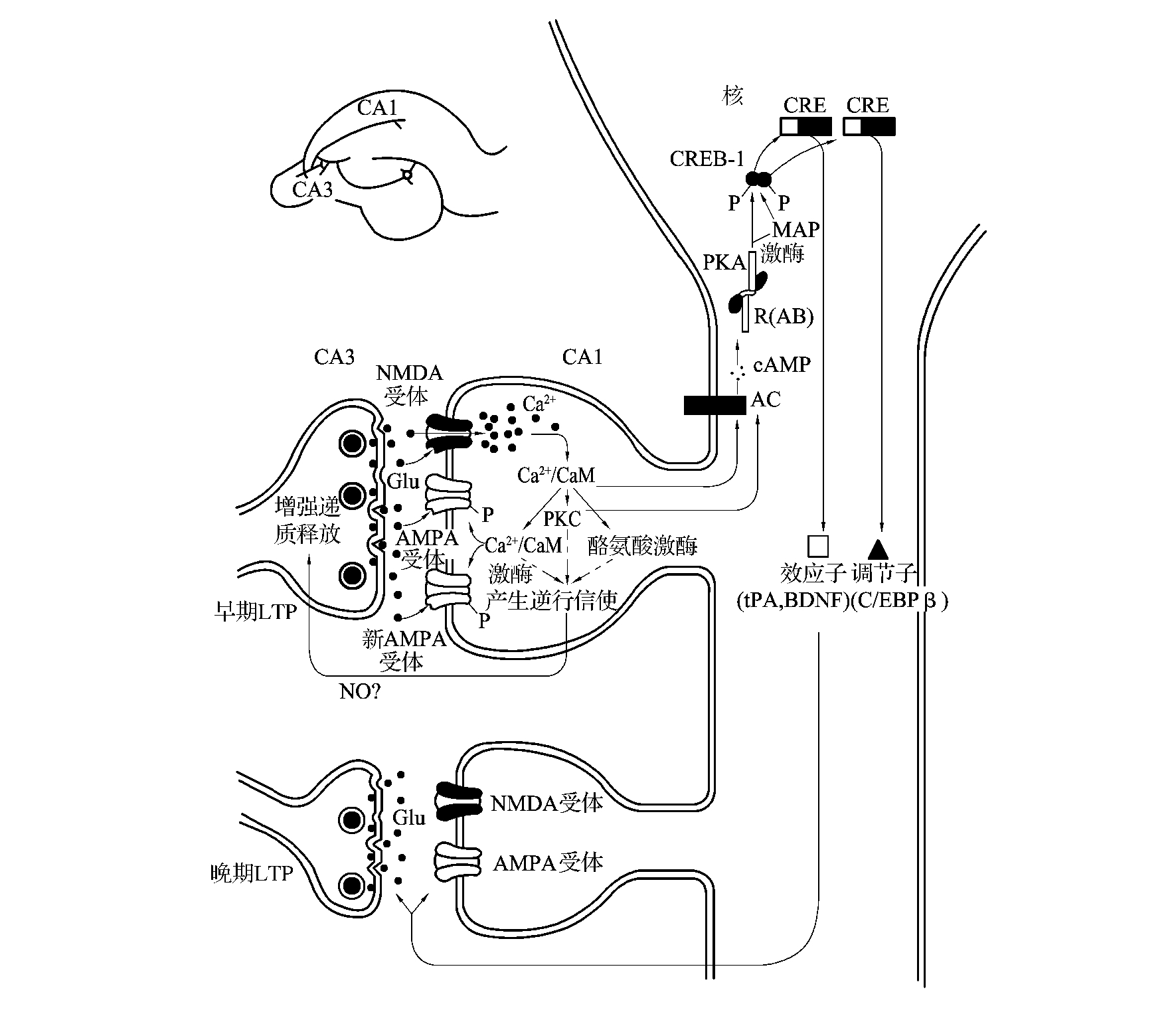
图4-3-8 早期和晚期长时程增强模式
可见长时程记忆与脑内的物质代谢有关,尤其是与脑内蛋白质的合成有关,而永久性记忆可能与建立新的突触联系及脑的形态学改变有关。研究表明,晚期LTP伴随着CA3细胞与CA1细胞之间新突触的生长。
(2)神经生理学机制
1)环路式突触联系:感觉性记忆产生的结构基础是神经元环路式联系方式,当各种感觉信息沿不同的途径传入中枢后,引起与学习和记忆相关脑区的大量神经元同时连续活动。在刺激停止后,神经元活动仍能继续一段时间,即出现神经元活动的后发放。(https://www.daowen.com)
2)海马环路:第一级记忆的维持及其向第二级记忆转化均与海马环路中神经的连续性活动有关。海马回路的神经元间形成的环路联系,可使该传入信息在环路中持续较长的时间,形成记忆的保持。与近期记忆有密切关系的神经结构是海马回路:海马→穹隆→下丘脑乳头体→丘脑前核→扣带回→海马。临床发现海马及其邻近结构损伤会引起近期记忆丧失。
3)突触的可塑性改变:长时程记忆的重要神经生理学基础是突触的可塑性改变,它主要来自于对习惯化、敏感化和长时程增强等突触传递现象的研究。而习惯化和敏感化的突触机制最早是在对低等海洋动物海兔缩鳃反射的研究中得以阐明的。
(3)神经生化机制与神经递质长时程记忆与脑内蛋白质的合成代谢有关。如果在学习过程中给动物蛋白合成抑制剂,长时程记忆会特异的丧失,而短时程记忆不受影响。可见长时程记忆需要脑内合成新的蛋白质。许多实验结果证实,中枢内有多种神经递质可以调节学习记忆过程。胆碱能系统与记忆有关,如果学习后立即给动物胆碱酯酶抑制剂—毒扁豆碱,动物记忆力明显提高。如果隔一段时间才给药,动物记忆力没有明显变化。临床上已观察到阿尔茨海默病是大脑皮层下区域的下橄榄核胆碱能神经元存在缺陷,导致学习记忆等认知功能明显下降,用胆碱酯酶抑制剂可治疗该病。胆碱能受体激动药可增强记忆,胆碱能受体阻断药则使学习记忆功能减弱。其次,去甲肾上腺素也与学习记忆有关。用利血平耗竭儿茶酚胺后,可破坏学习记忆过程。另外,血管升压素也可增强记忆,用该药治疗遗忘症在临床上已收到一定疗效。相反,催产素和脑啡肽可使动物学习记忆功能减退。
(4)神经解剖学机制新的突触联系的建立被认为是长时性记忆的解剖学基础。当储存记忆的皮层神经元受损伤时,如阿尔茨海默病,记忆无处储存,因而出现记忆缺陷。动物实验可见,生活在复杂环境中的大鼠皮层厚度大,突触联系多。反之则较少。人类第三级记忆的机制可能与此有关。
(二)生理性和病理性遗忘
遗忘是指部分或完全失去回忆和再认的能力,包括生理性遗忘和病理性遗忘。生理性遗忘是一种正常的生理现象,在学习后即刻开始,在感觉性记忆和第一级记忆阶段,遗忘的速率很高。产生遗忘的原因,主要是条件刺激长时间不能得到强化而引起消退抑制,或是后来信息干扰造成的。
病理性遗忘是指疾病情况下发生的遗忘称为记忆缺失或遗忘症。患者储存新的信息或回忆原有信息产生困难,在脑认知运动障碍的同时也常伴有记忆功能障碍,患者也可出现记忆丧失。患者对于过去的事情(尤其是其本人的经历)丧失记忆,在很多神经和精神病中常伴有记忆功能的缺损。
临床上把记忆障碍分为顺行性遗忘症和逆行性遗忘症。
1.顺行性遗忘症 凡患者不能保留新近获得的信息的称为顺行性遗忘症。临床发现大脑皮层联络区是记忆痕迹最后储存的区域,破坏联络区不同部位可引起遗忘症,海马和颞叶皮层损伤是造成记忆障碍的主要原因。临床上多见于慢性酒精中毒者、科萨科夫综合征(Korsakoff's syndrome)和阿尔茨海默病。内侧颞叶损伤的患者对新信息的获得能力发生障碍,而长时程记忆的回忆能力则不受损伤。
Korsakoff综合征的典型特征是顺行性遗忘,患者表现为长时程记忆的严重障碍,而短时程记忆的能力仍保持。该病患者死亡后病理解剖发现,海马、颞叶乳头体等的脑部有病理改变。AD患者有大脑皮层下区域的下橄榄核神经元缺陷、海马和颞叶联合皮层部位的损伤。
其发生机制可能由于信息不能从第一记忆转入第二记忆。由此可见记忆障碍的原因是多方面的,所以改善记忆的措施应是多种多样的。
2.逆行性遗忘症 表现为患者不能回忆脑功能障碍前一段时间内的经历,但新进入的信息仍能转入长时性记忆之中,第三级记忆也不受影响。其发病机制:可能是第二级记忆中的信息不能读出或第二级记忆本身发生紊乱。患者多见于脑震荡、电击和麻醉等。
二、语言和其他认知功能
(一)优势半球和皮层功能
语言是人类特有的一种极其复杂的高级神经活动,是随着人脑的进化发展而产生和完善的。人类进行条件反射的第二信号系统正是以语言功能为基础进行的。与语言有关的脑区位于大脑裂附近,人类大脑半球的结构和功能是不对称的,高级功能向一侧半球集中的现象称为一侧优势现象。①左侧大脑皮层在语言活动上占优势的现象称为优势半球。多数人语言功能定位于大脑左半球,理解和表达能力定位于左半球大脑皮层的不同区域。一侧优势现象虽与遗传有一定关系,但主要在后天实践中逐步形成,这与人类习惯使用右手有关,人类的语言左侧优势自10~12岁起逐步建立,此前如发生左半球损伤,尚有可能在右半球建立语言中枢。左半球在成年后不同区域损伤,很难在右半球建立新的语言中枢,将导致不同形式的失语。②右侧半球在非语词性认知功能上占优势,如对空间的辨认、深度直觉、触—压觉认识、图像视觉认识等。③两侧大脑半球对不同认知功能的互补性专门化现象,通过两侧大脑皮层的(胼胝体最大的连合纤维)连合纤维,将两侧皮层的功能联系起来,完成双侧的运动、感觉和视觉的协调。
(二)大脑皮层的语言中枢
Wernicke和Geschwind提出,当人们看到某一物体后到能说出其名称时的语言信息传导途径为:视觉信息由视觉感受器(视网膜感光细胞兴奋)传至外侧膝状体,经初级视皮层(17区)到达高级视皮层(18区),经顶—颞—枕叶的联合皮质—角回(39区),该区与听、视、体感信息联合有关;从此处投射至颞上回后端的Wernicke语言区(22区,又称后说话区),使视觉信息转化为该词的语声表象,一旦声音模式形成,经该区的纤维通过弓状束在传到中央前回底部前方的Broca区(45区,又称前说话区),将传来的信息处理为相应的发声形式,进而传至运动皮层支配面部的区域(4区),启动唇、舌、喉的运动而发声,说出该物体的名称。这一信号传递途径如图4-3-9中所示。若要求受试者完成复述一个听到的词语时,其听觉信息传导通路除经听神经传至内侧膝状体,经初级听皮层(41区)传至高级听皮层(42区)外,在角回以后的传导通路与视觉信息传递途径相同。因此,他们提出表达视觉和听觉信息的Wernicke-Geschwind模型,并在临床上得到广泛应用。
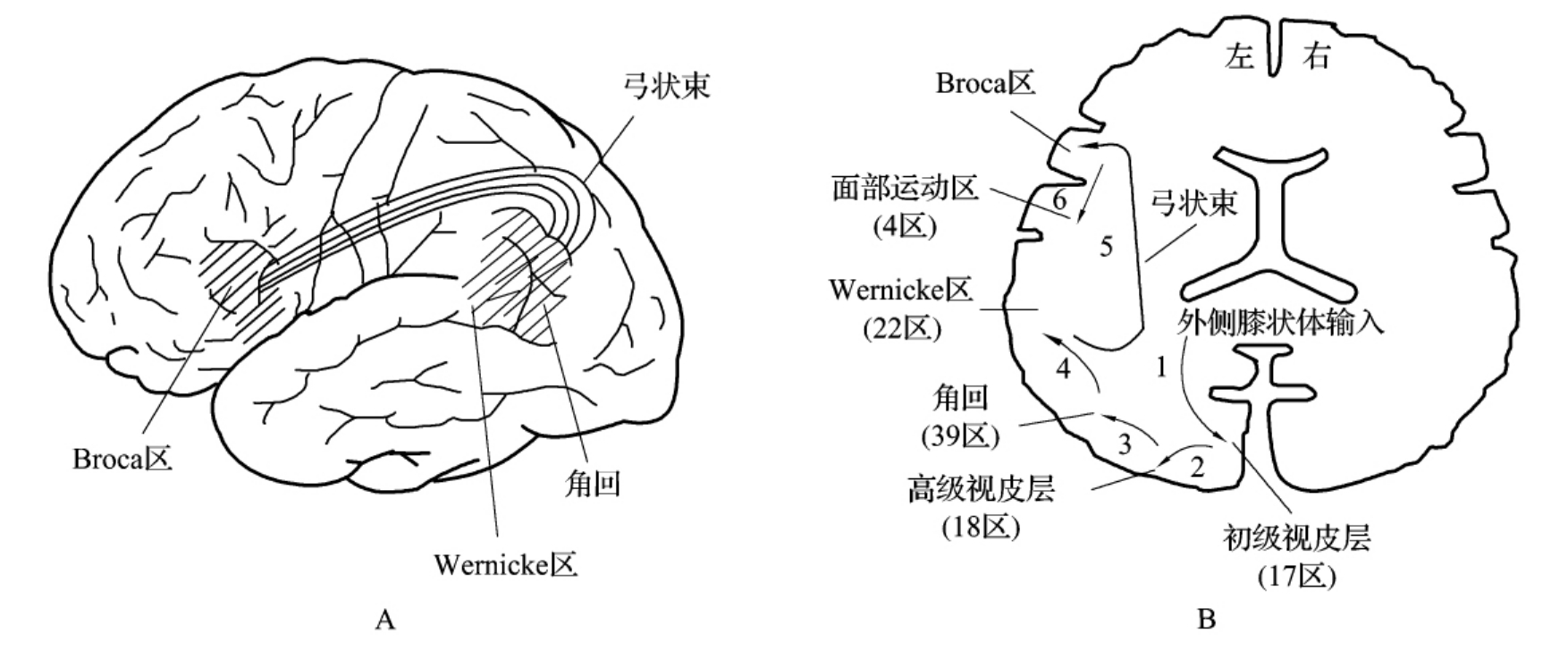
图4-3-9 A.语言功能有关的脑区部位和纤维联系;B.看到某一物体后到能说出其名称时的语言信息传导途径
根据Wernicke-Geschwind模型可预示不同部位病变所产生的不同失语症,助于理解失语症的症状及类型。但近年研究特别是PET观察发现,该模型尚有以下不足之处:①该模型强调Wernicke区和Broca区在语言接收与表达方式方面的重要性,但单纯某一区损伤的典型症状通常是周围区损伤的结果;以健康志愿者为研究对象,对语言功能脑机制的研究验证了这一现实。例如进一步用动词产生作业(见图4-3-9)要求受试者根据所呈现的名词说出相关动词时,可见激活的脑区有扣带回前部、左侧前额叶和额眶部、左侧颞叶后部、右侧小脑。其中左侧前额叶和左侧颞叶后部分别相当于Broca区和Wernicke区。②该模型过分强调皮层区域的重要性,而某些皮层下结构,如左侧丘脑、左侧尾核及其邻近白质对语言功能也有重要作用;例如仅呈现给受试者一个“十”字和一系列英语单词(图4-3-10),不要求作任何反应,分析和确定言语所依赖的视觉系统功能,发现视觉加工时,左侧纹状体外皮质的活动明显,说明左侧纹状体外皮质在语言中具有重要作用。③并非所有的听觉传入都是以同样的途径完成的,还有处理声音(语声学方面)和含意(语义学方面)相互分离的途径。例如用复述作业要求受试者(图4-24)复述所看到或听到的单词时,可见激活运动皮质、辅助运动区、岛叶和小脑中部等脑区,但没有见到激活经典的语言功能区Broca区和Wernicke区。
(三)语言的中枢功能定位与神经过程的近代模式
音、形、义既是文字的三要素,也是语言加工的三个功能系统。①语音加工的中枢功能定位:听单词则可激活左侧额叶上回和额叶中回的后部。提示该部位是语音感知的脑功能区。对单词和音节的识别,主要激活前额叶特别是额下回的后部。②字型加工的中枢功能定位:fM IR研究发现,对字母的加工和对线条的加工可能有着本质的不同,真词和伪词加工的过程所依赖的脑区也各不相同。③语义加工的中枢功能定位:语义加工只激活左侧额叶后部。颞叶可能是语义的存储地,额叶则主要是控制和输出的部位。
近代对正常人和失语症病例的认识和fM IR研究结果表明,语言的中枢功能定位于大脑皮层的不同区域,并具有复杂的相互联系,近代语言神经过程的模式,较Wernicke-Geschwind模式更复杂,如图4-3-10所示。
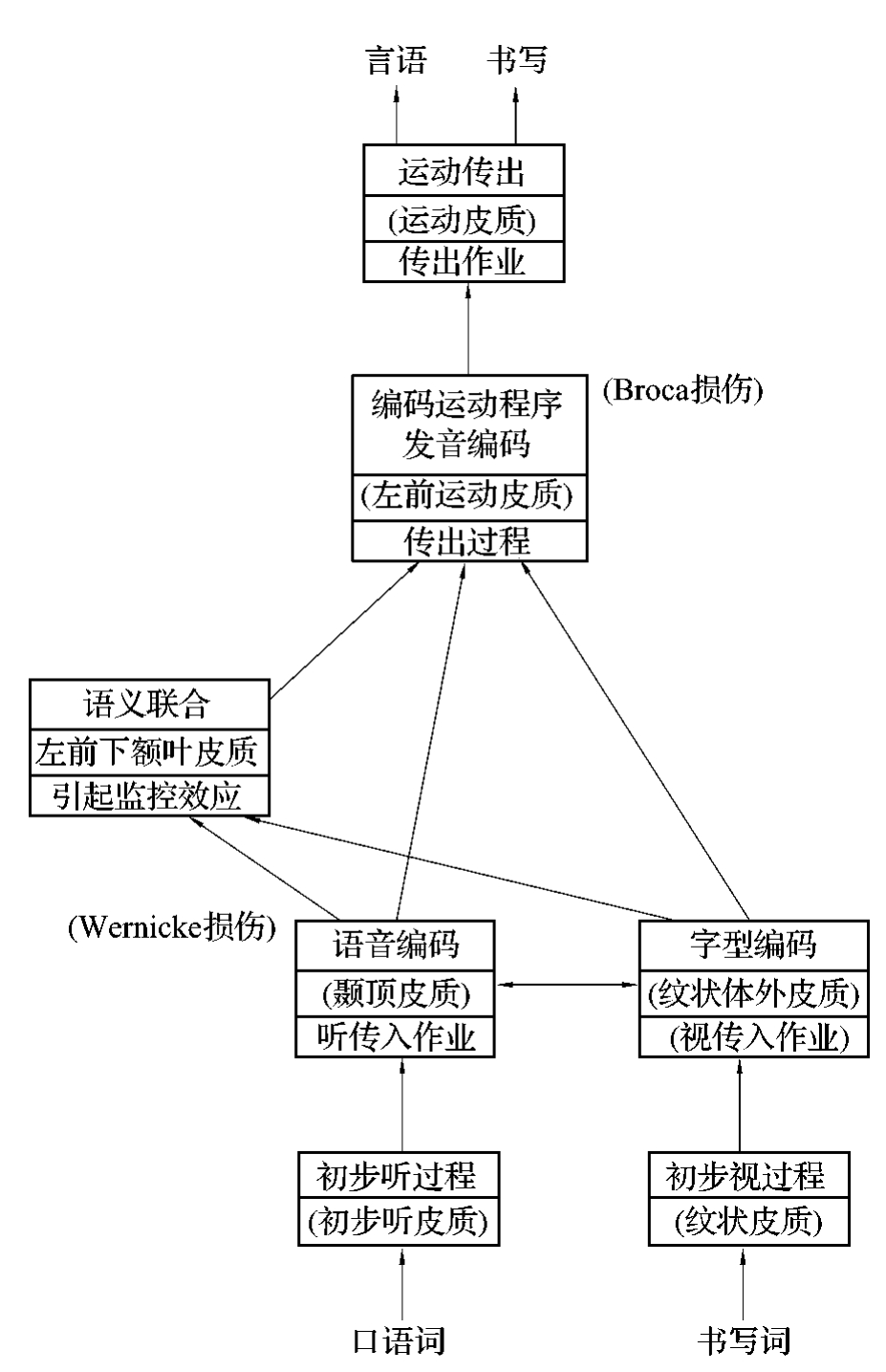
图4-3-10 语言神经过程的近代模式
(四)语言功能障碍
AD患者的主要病变特点为大脑皮质、海马、某些皮质下神经核如杏仁核、前脑基底神经核和丘脑有大量的老年斑和神经元纤维缠结,特别是神经炎性老年斑,软脑膜和皮质血管壁都有β淀粉样蛋白沉积。这些病理改变导致语言优势侧大脑皮层特异区域的损伤,引起各种特殊形式的语言功能障碍。
AD患者的左侧丘脑损伤时,会出现类似跨皮层感觉性失语的症状,患者对口头语言的理解力差,而复述能力正常。AD患者的Wernicke区与后顶—颞联合皮层间的联系中断,常出现流畅性失语,即口语流利,但有明显的理解障碍和找词困难。患者不能读写,很难到适当词语,但能流畅地复述别人的话。AD患者的Broca区至辅助运动皮层之间的联系中断,损伤部位通常位于左额叶内侧部或边界区前部。由于边界区包括对词义记忆有重要功能的联合皮层及对技巧运动有重要作用的辅助运动皮层,患者复述、理解能力的损伤较轻,而书写则有严重障碍。AD患者的大脑皮层外侧裂周围区域,同时Wernicke区、弓状束和Broca区也有损伤,可致患者完全性失语常伴有完全性右侧偏瘫和右侧感觉障碍以及右侧同侧偏盲。若AD患者的脑部损伤的部位是连接Broca区和Wernicke区的弓状束,则造成两区的功能分离。患者复述、命名严重受损,语言流畅,但有许多言语错乱表现,常将错误的词或声音替换正确的词或声音。患者能默读并理解文意,书写时可能出现一定程度的随意运动障碍。在AD患者额叶的运动联合皮层损伤时,一般还涉及44区和45区,即额叶盖(Broca区)的一部分。重症患者还有运动前区和前额叶(6、8、9和46区)的损伤。患者可以看懂文字,听懂别人的谈话,但自己却不会说话,不能用词语作口头表达,只使用关键词,也无法将几个短语组分协调地构成一个句子,复述能力受损,但对口头和书写语言的理解是正常的。损伤主要涉及左侧颞叶后部或Wernicke区(Brodmann 22区),但常延及颞叶上部的40区、39区以及颞叶下部的37区。其特征是对语言的理解力发生明显的缺陷。患者可出现言语错乱、语言过多或空洞,但对语言的理解受损,复述单词和短语的能力也受损。
由此说明,完整的语言功能与大脑皮层的广大区域的神经活动有关,各区功能之间密切相关。用PET和fMRI研究语言障碍以及相关的疾病,发现进入脑的信息在引起语言活动前,并非只有一条传输通路传送信息。PET研究发现,当看见画面上的字和听到词的发音时脑的活动部位不同,当听到发音时,Wernicke区处于活动状态,当字只看见而未听到词的发音时,Wernicke区就不活动。视觉信息可以直接送到Broca区不需先送到Wernicke区就能直接讲出来。这说明看字和听词的语言通路不通。如果既没看字也没听词,只是在想一个词的意义时,激活了大脑的广泛区域,但是部位不同。因此认为,大脑对语言的处理过程呈并联通路和串联通路的复杂网络连接。
三、视空间功能与视空间功能障碍
视觉是表象最丰富的感知形式,视觉空间认知功能包括物与物、物与观察者、景物之间的空间方位关系。视知觉的形成过程是视网膜接受视觉刺激后,物像经过视网膜处理,形成不同的“像素”,经过视系统的传导通路到达中枢,视觉信息在超过30个功能性皮层区中进行处理,视皮层区有物体和空间两条处理通路,都投射到额叶的部位,额叶也发送反馈信息回到视皮层处理区。有人认为这种反馈的投射在视觉中起自上而下的作用,比如分配“注意”到所选择的视觉刺激。视皮层处理区把由不同神经通路所传递的关于颜色、运动、形状、亮度的复杂信息进行平行分析、加工和处理,最后成为综合知觉。这一过程中,必须有选释性“注意”的参与,通过对周边感受器所接受的感觉信息的过滤、筛选,再通过对信息进行取舍,突出对生存和生活有重要意义的视觉目标。经过注意过程处理后的视区间的相互关系就形成了对该视觉目标感知的基础,最终形成综合的视知觉。
(一)视觉中枢的神经机制
1.中枢视觉通路
(1)大细胞通路(magnocellular pathway,简称M-通道)起源于视网膜α型神经节细胞(M-细胞),其轴突纤维投射到外膝状体大细胞层(M层),再投射到纹外皮质(初级视皮质的4Cα),然后通过V2区(Brodmarn 18区,次级视皮质)的粗条投射到Vs(中颞区,middle temporal,简称MT区、Brodmarn 19区、高级视皮质),MT区能够被运动刺激选择性的激活。有人发现连续运动、光流、生物体运动三种刺激,每一种都激活了MT区的特定部分。另一条支路是从外膝状体的M细胞经4B中的方位选择性细胞,再直接或通过V2的粗条投射到V3,后者是所谓的动态图形识别通道。M-通道最后到大脑的背侧、更高级的视皮质的联合区,包括内上颞区(medial superior temporal,简称MST),腹内顶区(ventral intraparieta,简称VIP)和7a区等,所以称为顶背侧通路。
(2)小细胞通路(parvocellular pathway,简称P-通路)起源于视网膜P型神经节细胞(P-细胞),其纤维投射到外膝状体的小细胞层(P-层);在投射到皮质V1区的4Cβ,从这里一条通过2、3层内的斑点,V2的细条到V4(属Brodmarn 19区是高级视皮质),是主要的色觉通道;另一条通过斑点间和V2的粗条到V4,与图形识别有关。P-通道最后到大脑腹侧的下颞区(inferior-temporal,简称IT区),所以称为腹侧通路。
2.视觉通路的中转站视网膜神经节细胞层兴奋的空间模式点对点的地投射在外侧膝状体、上丘和视皮质。
(1)外侧膝状体 人外侧膝状体分为六层,每一层有两类细胞,一是主细胞投射至皮质,二是局部的中间神经元,视神经纤维和初级视皮质的细纤维,与外膝体各层的主细胞、中间神经元的树突形成突触。
(2)初级视皮质 又称纹状皮质,位于大脑枕叶,接受来自外侧膝状体主细胞的轴突,形成视放射,也和其他皮质区域有广泛地连接。初级视通路在视觉功能中起重要作用,若外侧膝状体—皮质通路损伤,将引起视功能丧失。
(3)纹状外视区 纹状皮质以外的视皮质区域(如Brodmarn18、19区等)的损伤也可导致视功能的紊乱,这样的患者不能正确利用初级视通路所提供的视信息,难以辨认物体、图像或颜色,常伴有语言、阅读、书写困难。
3.人视觉的脑功能定位 视觉功能区域可分为两类通路:①腹侧通路与视觉特征检测和物体辨认有关,大多位于皮质的腹侧;②背侧通路主要负责物体空间位置和运动方向的识别,大多位于皮质背侧。位于腹侧的枕、额区主要负责颜色、形状、纹理等特征的知觉和物体的识别,而空间和运动知觉则主要依赖于枕叶的背侧和顶叶皮质。参与颜色知觉的功能区位于枕叶的腹侧,视觉系统对颜色的加工有一定的层次性。枕叶外侧部MT区只对具体的照片或图片刺激起反应,但对熟悉程度不同的刺激物(如名人脸孔、艺术、雕塑等)的反应却没有差异,推测该区可能是一个从初级到高级加工的中间环节。参与脸孔识别有关的脑区,从后到前其功能的复杂程度是逐渐递增的,如辨别脸孔的性别主要激活腹侧额叶的后部,而识别具体的脸则激活腹侧额叶的前部,若进一步说出职业,所激活的脑区就移至颞叶的最前端。Peterson等发现文字刺激主要激活左侧纹外皮质的内侧部认为在视觉的腹侧通路可能还包括含感知文字的专门功能区域。
注意刺激物的颜色和运动特征可分别激活MT区。利用fMRI观察发现,注意运动光点比注意静止光点可更显著地激活MT区。Fink等发现使受试者注意局部(小字母)主要激活左侧枕叶下部,而注意整体和注意局部的脑机制不同。Shulman等对已发表的有关脑功能成像研究报道进行综合分析后发现,在大多数实验条件下注意对初级皮质具有一定调节作用。
(二)视空间功能障碍
由视觉原因造成物体在空间内的各种特性的认知障碍称为视觉性空间知觉障碍,简称视空间功能障碍。主要表现在四个方面:视觉定向力障碍、视觉立体障碍、半侧空间知觉障碍和地方判断力障碍。在AD早期即可出现视空间功能损害,一般认为视空间功能障碍的病灶主要在顶、枕、颞叶的交界处,以右半球(非优势半球)顶叶病变为主。中、重度AD患者视空间功能损害变化的总趋势是在每个阶段均明显下降,与AD患者病程特点的总趋势相吻合,这可能与大脑皮层主管视空间功能相应脑区萎缩病变的范围及程度逐渐增加有关。Johnson等实验表明,AD患者在作类似的角度辨别时背侧通路的顶上小叶激活明显减弱,而腹侧通路上的梭状回代偿性增强。
(周玉琴 张孟业)
参考文献
1.Bubber P,Haroutumian V,Fish G,et al.Mitochondrial abnormalities in Alzheimer brain:mechanistic implications[J].Ann Neurol,2005,57(5):695-703
2.丁绍红,施建华,尹晓敏,等.PKA、GSK-3β和cdk5对微管相关蛋白tau的位点特异性磷酸化[J].南通大学学报2006,26(4):235-238
3.王琳,王晔,郭大文,等.Alzheimer病患者脑脊液tau蛋白水平的检测及临床意义[J].临床神经病学杂志,2005,15(3):352-353
4.肖岚,李巍,蔡文琴.芳香化酶在神经干细胞及其分化后细胞中的表达.解剖学报.2000,31:118-121
5.Tomoko Ishibashi,Kelly A Dakin,Bet hstvens,et al.Ast rocytes promotemyelination in response to elect rical impulse[J].Neu2 ron,2006,49:823-832
6.DiPatre P L,Gelman B B.Microglial cell activation in aging and Alzhe-imer disease:Partial linkage with neurofibrillary tangle burden in the hippocampus[J].J Neuropathol Exp Neurol,1997,56: 143-149
7.Roskoden T,Linke R,Schwegler H.Transient early postnatal corticosterone treatment of rats leads to accelerated aquisition of a spatial radialmaze task and morphological changes in the septohippocampal region[J].Behav Brain Res,2005,157(1):45-53
8.German DC,Yazdani U,Speciale SG,et al.Cholinergic neuropathology in amousemodel of Alzheimer's disease[J].JComp Neurol,2003,462(4):371-381
9.M aslioh E.Deficientglutamate transport isas2sociated with neurodegeneration in Alzheimer's diease[J].A nnal of N euro logy,1996,40(5):759-766
10.Ferrerl,Marti E,Tortosa A,et al,Dystrophic neunites of senile plaques are defective in protein involved in exocytosis and neurotiansmission.JNeuropathol Exp Neuo1,1998,57(3):218-225
11.Ballard C,JohnsonM,PiggottM,et al.A positive association be2 tween 52HT re2up take binding sites and dep ression in dementia with Lewy bodies.Affect Disord,2002,69:219-223
12.Uzakov S,Frey JU,KorzV.Reinforcement of rat hippocampal LTP by holeboard training.LearnMem,2005,12:165-171
13.姚泰主编.8年制临床医学专业用生理学.北京:人民卫生出版社.2005
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。



