在生物演化过程中,结构与功能的演化是同步的。在种系发生上,人脑的高度发达,使管理各器官功能的高级中枢——大脑皮质高度分化。1861年,Broca观察了出生前有语言缺陷的人脑,发现损伤部位在额下回。Jackson于1864年发现刺激中央前回,可引起部分性癫痫。后人根据大脑皮质的结构与功能将其分为躯体运动、躯体感觉、视觉和听觉等区。亦有人把功能相对明确的部位称为中枢(center),功能相对不明确的称为哑区(silentarea)。此后逐渐得知各功能的主要投射部位为中心区,其周围的皮质称为边周区。边周区与中心区结构相似,功能也受其控制。各边周区之间的脑区称联络区(associational area),它们多集中在额叶前部,顶、枕叶的中部和颞叶的中、下部等处。现认为这些在种系发生上出现较晚的区域,联系广泛,是更高级的皮质,很可能具有联络和整合的功能。
从形态上,人们以中央沟为界将大脑皮质分为前、后二部,前部与运动有关,后部与感觉有关。但从功能上,如此机械分区是不确切的。中央沟之前除有第Ⅰ躯体运动区、第Ⅱ躯体运动区、附加运动区,以运动为主之外,也接受少量的传入冲动;中央沟之后除有第Ⅰ躯体感觉区、第Ⅱ躯体感觉区等,以感觉为主而外,也发少量纤维参与运动反应。因此有人称中央后回为感觉运动区(sensorimotor area)[5]。
大脑皮质全部或大部都有传入和传出的特点,只是人们通常把功能上以传出为主,接受为辅的区域,称为运动区(motor area);以接受为主,传出为辅的区域,称为感觉区(sensory area);其间广大的区域即所谓的“联络区”(图3-2-1)。
下面以躯体运动区、躯体感觉区、视区、听区、语言区、内脏脑和脑病的临床表现来叙述脑的功能分区。
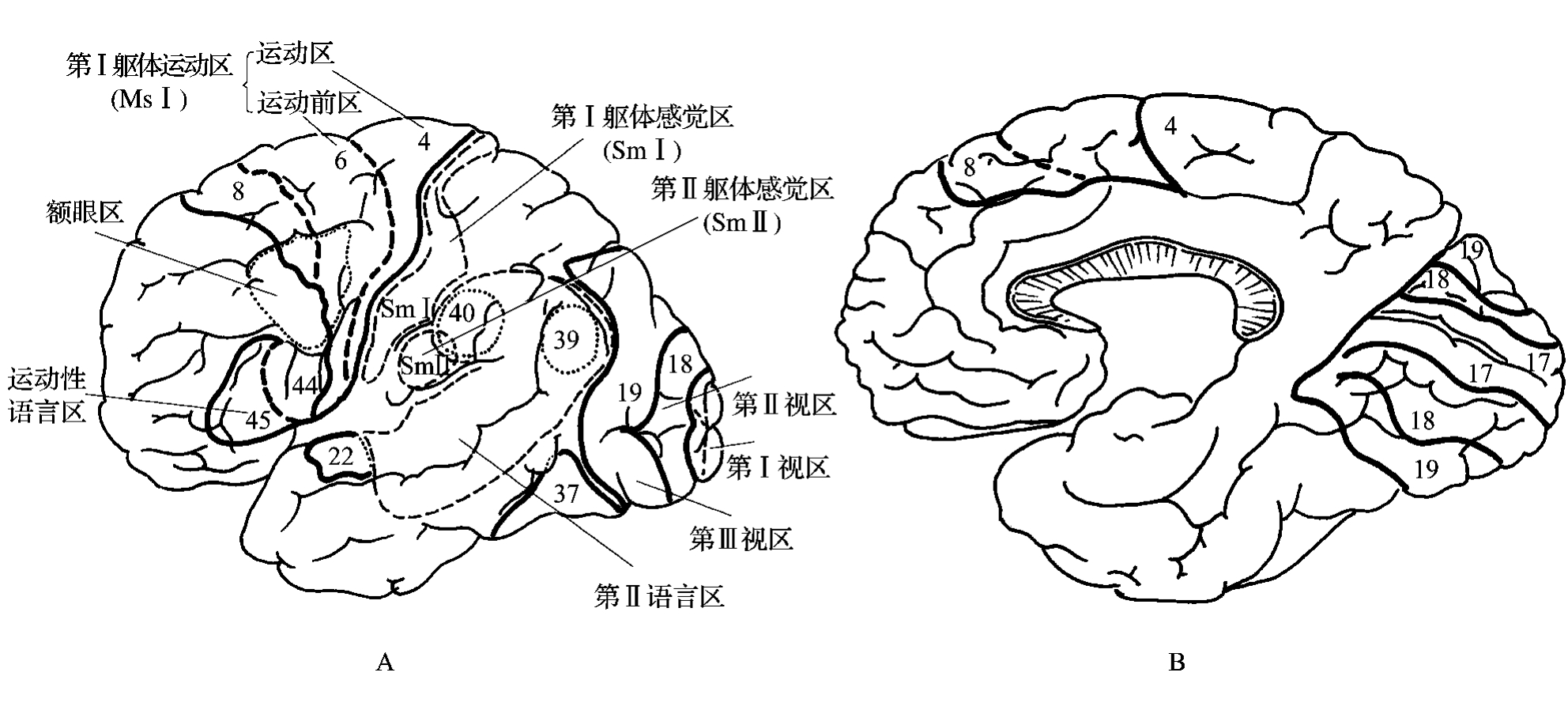
图3-2-1 皮质功能区划分示意图。A.上外侧面;B.内侧面
一、躯体运动区
躯体运动区是指与躯体运动有关的脑区,主要是额叶皮质。人类额叶皮质高度发达,尤其是前额区最为明显,且随种系的进化而增加。猫的额叶皮质仅占全部皮质的3%,黑猩猩占16.9%,而人则高达29%。额叶的功能与躯体运动、头眼运动、发音与语言以及高级思维活动有关,也是情绪活动的重要中枢[6]。
根据额叶细胞构筑、纤维联系和功能,通常分为两个区,即中央前区和前额区。前者大部分是运动区,后者属于“联络区”。
(一)中央前区
中央前区(precentral area)包括中央前回、中央旁小叶前部和额上、中、下回的后部。中央前区又分为前、后两部。后部称运动区,相当于Brodmann 4区,它上宽下窄,位于中央沟前方,占中央前回的大部分,在大脑内侧面占中央旁小叶前部,至额下回水平,仅占中央沟前壁。前部称运动前区,各家所指范围有所不同,包括额内侧回、额上回和额中回的后部,全区也是上宽下窄,下部最窄处仅限于中央前回。运动前区相当于Brodmann 6区。6区通常又分6aα、6aβ和6b三区(也有人命名为6p、6a和6b)。除6区以外,有人主张运动前区还包括8区、44区和45区的一部分。
中央前区细胞的结构特点是:①皮质极厚,4区是全部大脑皮质中最厚的区域。②属于无颗粒皮质,几乎没有颗粒细胞,因此第Ⅱ层不发达,没有第Ⅳ层,且各层很难区分。③第Ⅲ层很厚,又可分为三个亚层,有大型锥体细胞。④第Ⅴ层最厚,由锥体细胞组成。4区此层有三五成群的Betz细胞。此细胞是巨型锥体细胞,高60~120 um,宽15~70 um,中央旁小叶者最大,中央前回下部者最小。人的Betz细胞总数约3.4万个,但分布并不平均,75%集中于4区的下肢代表区,18%集中于上肢代表区,7%集中于面代表区。⑤第Ⅵ层也较厚,但与白质交界不明显。⑥细胞在纤维之间排列成柱状,在切面上呈放射状,称柱状结构。
Betz细胞的轴突大部分组成皮质脊髓束。皮质核束的纤维可能来自较小的锥体细胞。锥体束的纤维并非全部始自4区,始自4区者仅占31%,始自6区者占29%,还有40%来自顶叶的3、1、2、5和7区。锥体束纤维的直径也大小不一。1~4 um者占90%,它们来自上述各部第Ⅲ、Ⅴ层的锥体细胞;11~22 um者仅占2%~3%,它们来自4区第Ⅴ层的Betz细胞。
1.第Ⅰ躯体运动区(primary or first somatomotor area,Msl)此区范围各家有所出入,本文仅指Brodmann 4区。有人主张还包括6aα区,有人主张将整个中央前区属之,但以4区为主。从比较解剖学的角度观察,此区随着进化越来越小(图3-2-2)。
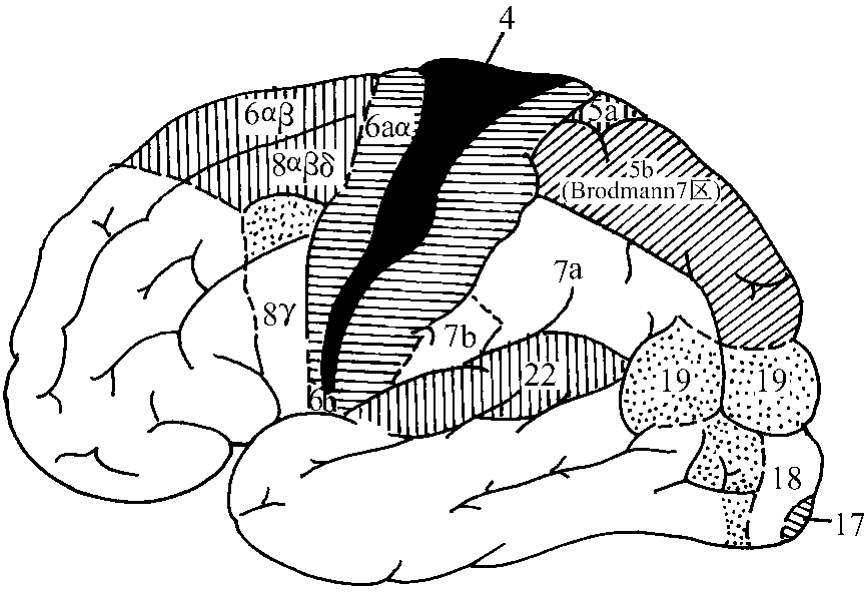
图3-2-2 人脑电刺激能诱发运动的皮质区
4区的传入纤维有:①经丘脑腹外侧核中继的源于小脑的纤维;②来自丘脑、下丘脑的其他感觉性纤维;③来自1、5、7、8、9、10区的联络纤维;④经胼胝体来自对侧4区的连合纤维。这些传入纤维与一些锥体细胞构成垂直联系,通过颗粒细胞又形成水平联系,这样来自多方面的冲动很容易使一些锥体细胞兴奋。4区的传出纤维主要管理对侧半身的躯体运动。主要纤维是锥体系,此外还有锥体外系、皮质丘脑下丘脑纤维、皮质脑桥纤维和皮质网状纤维[7]。4区的功能特点如下。
(1)支配对侧。4区的Betz细胞及其他锥体细胞发出纤维构成锥体束,这些纤维通常交叉至对侧,控制对侧一个或一组骨骼肌。但该交叉并不完全,多数头面肌(面下部肌和颏舌肌除外),咽、喉肌和躯干肌等受双侧支配,司双侧联合运动。因此一侧锥体束受损,这些部位的骨骼肌并不瘫痪,症状不明显。
(2)具有精确的机能定位,且有一定的规律。自下而上依次是舌、颌、唇、喉(发音)、睑、额、颈、指(拇指最低,小指最高)、掌、腕、肘、肩、躯干、股、膝、踝、趾,形似倒置人形的投影。头面肌代表区在底部(头面肌之内部安排仍取正位),上肢代表区在中间部,下肢代表区在上部(小腿、足的代表区在半球内侧面的中央旁小叶前部)。此外,中央旁小叶前部还有肛门外括约肌和膀胱外括约肌的代表区。4区损伤,只导致肢体或颜面有关部位骨骼肌瘫痪,但粗大的联合运动经过一定时期可以恢复,而精细和单独的运动难以恢复。
(3)机能代表区的大小与运动的精细复杂程度有关。运动愈精细复杂的部位,其代表区愈大。例如手与五指所占区域几乎等于整个下肢代表区。而手代表区,又以拇指为大(图3-2-3)。
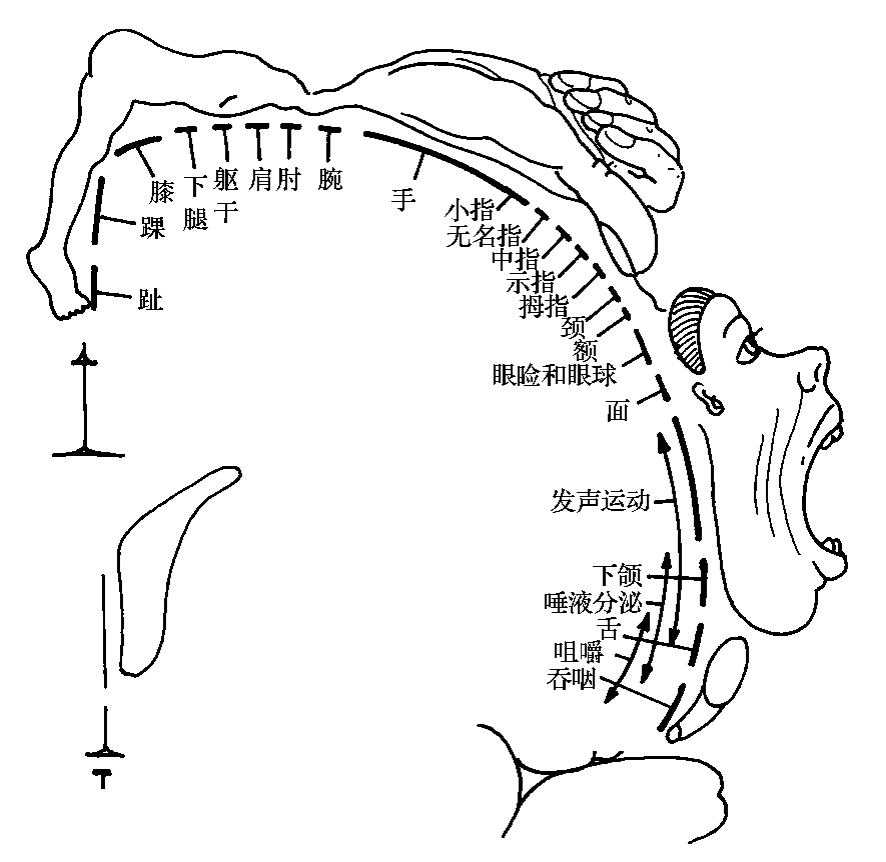
图3-2-3 第Ⅰ躯体运动区功能定位示意图
(4)刺激代表区所产生的运动相当简单,主要是单块骨骼肌的收缩,甚至是某块肌的某一部分收缩,因此不发生肌群的协同收缩。如此运动的结果,只能是单关节或多关节的运动,如新生儿那样,不能司理技巧性运动。
2.运动前区(premotor area)本书指Brodmann 6区。在个体发生上,6区的发展比4区快,至成人占全部脑皮质的7.5%。皮质结构与4区相似,主要由大锥体细胞组成,但无Betz细胞。6aα区位于4区的前方;6aβ区位于额上回,其内侧面的部分是附加运动区的一部分;6b区最小,位于面区的前方。
6区的传入纤维有来自丘脑腹前核和腹外侧核的投射纤维,同侧半球4、1、5、39、40、42等区的联络纤维,对侧半球6、4、1、5、39等区的连合纤维。
6区是躯体运动的重要起点之一。电刺激6aα区与电刺激4区引起的反应相似,但需要的电流较强。切除4区后,再刺激6区,不能引起单个运动,这说明运动前区的冲动先传至4区,再经皮质脊髓束到达运动神经元。强烈刺激6aβ区,引起头、眼、躯干转向对侧以及对侧肢体屈或伸的协同运动。此运动与4区无关,切除4区仍可引起。刺激6b区,引起面肌、咀嚼肌和咽喉肌的协调运动。如一侧6b区受损,运动缺陷较轻,或无运动缺陷。若切除一侧6区和4区,对侧身体产生痉挛性瘫痪;若只切除6区以及4区和6区间的窄条皮质区(曾称为条区strip area,简写为4s区),也可产生痉挛性瘫痪。若只切除猴的4s区,可引起对侧暂时性轻瘫和永久性的伸肌张力亢进;若切除4区的后部,只引起对侧弛缓性轻瘫。以上说明痉挛性偏瘫主要与6区有关。
3.额眼区(frontal eye field)位于额中回的后部,相当于6、8、9区的相邻部分,位于面区与手区之间。它是与眼肌随意运动有关的皮质区,此外与眼肌运动相关的头颈肌的协同运动也有关系。人脑的额眼区,司两眼的同向偏斜,一般偏向对侧。与此同时头颈肌有相应的协同运动,例如眼转向一侧,头面也转向同一侧。此外,刺激额眼区可引起瞳孔扩大。眼的随意运动和反射运动,无疑是通过额眼区及其投射纤维完成的。现已清楚,额眼区发出纤维经皮质核束下行,多数纤维经网状结构中继,止于眼肌诸核,只有少数纤维直接止于眼肌诸核。此外,还可能经基底核中继后再下行至中脑被盖网状结构,联系眼肌诸核。视觉引起眼运动的径路是视觉冲动通过17区,经枕额下束达额眼区。此外,额眼区与丘脑背内侧核有往返联系。额眼区损伤,两眼转向瘫痪侧,此种转动一般是暂时的。
4.附加运动区(supplementary motor area)相当于半球内侧面6区和8区的一部分。有人认为此区很小,只占6aβ的一部分,直径仅1cm。此区也属感觉运动区,有人把它称为第Ⅱ躯体运动区(MsⅡ),有人把它称为第Ⅲ躯体感觉区(SmⅢ),但与发音以及说话也有一定的关系(见下文)。电刺激此部皮质,可引起身体各部的运动,只是刺激强度比第Ⅰ躯体运动区者为高。其局部定位在猴是:头部位于嘴侧,向后依次是前肢、后肢;背侧朝向胼胝体,四肢伸向半球的背内侧缘。人与猴的相当。人脑此区兴奋时,可引起对侧上肢的上举,头、眼的协同转动和两侧躯干肌、下肢肌的协同收缩。此外,还可引起瞳孔扩大、心跳加快、发音或说话遏止等。这些现象说明附加运动区既有躯体运动的功能,又有内脏运动的功能。切除中央前回,再刺激此区,仍有上述运动,可见此区是一个独立的功能区。
5.第二运动区(second motor area)人和猴都有第二运动区,它位于中央前、后回最下端(中央间回),并伸入外侧沟。通常此处中央沟已终止,中央前回与中央后回在此相移行。此区有双侧上、下肢的代表区,但无头部(面、舌、口、喉)的代表区。切除第Ⅰ躯体运动区后,再刺激第二运动区,对侧出现手、足运动,说明此区在功能上是一个独立的区。它发出的纤维大概也加入锥体束。实际上,电刺激人脑能引出运动的区域很多。除上述诸区外,还有3、l、2、5、19、22区,刺激之也能引起运动。
(二)前额区
前额区(prefrontal area)在中央前回之前,又叫额极区。此区在进化上较新,灵长类较发达,人类则特别发达,几乎相当于整个额叶。前额区的划分比较混乱,各家记述多不一致。本书此区包括额上、中、下回之大部以及眶回和额内侧回。该部分相当于Brodmann 8、9、10、11、12、32、44、45、46和47区。其中9、10、11、12、45、46和47区是固有前额区,而8和44区是前额区和中央前区的过渡区,32区是前额区和边缘叶的过渡区。眶回又叫眶额区(orbitofrotal area)。
人脑前额区高度发达,与各叶皮质均有往返联系,因此称联络区比较恰当。它通过上、下枕额束与同侧枕、顶和颞叶相联系;通过胼胝体与对侧前额区相连合,与对侧其他皮质区也有连合纤维;与丘脑、纹状体和下丘脑也有很多联系,其中9、10区和眶回及丘脑背内侧核之间往返纤维很丰富,眶回的纤维投射到下丘脑的旁正中诸核;前额区的皮质脑桥纤维比第Ⅰ躯体运动区的纤维为少,它们至脑桥核的旁正中部分;此外,此区纤维还可至脑干其他运动核以及小脑等处。
前额区的联系对躯体和内脏活动都有极复杂的影响,特别是眶回。刺激猴的眶回,可降低呼吸、血压和胃的蠕动,并抑制运动皮质和反射所引起的运动。切除灵长类的眶回,可引起运动过度或不稳定等现象,与切除扣带回前部的结果相似。眶回后部和扣带回前部皮质常被认为是边缘系的一部分,与呼吸、血压等自主神经系功能关系密切。
对前额区功能的了解,多来自前额叶切断术的患者。20世纪40年代,医学界曾盛行切断额叶的纤维联系以缓解精神病患者的顽固性疼痛,特别是切断8、9、10、11、12、45、46、47区与丘脑背内侧核间的往返联系。这种止痛作用也包括内脏痛。但止痛的同时患者的性格、行为和对近期的记忆也有改变,其结果是精神涣散、判断力差、智力减退和抽象思维能力降低。由此推测前额区与人的抽象思维和高级智力活动有关。
脑血管性痴呆的病理因素不仅在于梗死灶的大小,还取决于梗死部位、年龄等其他因素,梗死部位与痴呆程度的关系依次为额叶;纹状体前部;丘脑;枕叶,而海马、海马旁回和胼胝体前部的小梗死灶也易出现痴呆。
附:抑制区(suppressor area)
1936年,Hines发现猴脑4区之前6区之后的一个狭长区域(简称48区),刺激该区域可抑制伸肌的张力,切除该区域可出现暂时的痉挛性瘫痪,后来发现人也是如此,猩猩也有相似的区域;此外,8区、2区、19区、24区也可产生相似的抑制效应。当时人们称这些区域为“抑制区”。但是近来人们知道这种抑制是“扩散性抑制”扩散的结果,而与任何一个特定的皮质区无关。只是这样还不能圆满解释抑制何以长达数分钟,而且数分钟内,运动反应不是完全停止,就是完全变了样子。人们推测这一现象可能与新皮质内的一种细纤维(有时是单胺能的)的旁分泌效应(paracrine effect)有关。总之,近半个世纪的研究和争论,人们现在倾向于认为大脑皮质没有什么特定的专门的抑制区。它与大脑皮质活动的抑制是两个完全不同的概念,后者是一个正常的生理学过程。
二、躯体感觉区
躯体感觉区是指与躯体感觉有关的脑区,主要是顶叶皮质。顶叶皮质可分为三个区,即中央后区、顶上区和顶下区,与躯体感觉有关的主要是中央后区和顶上区两部分。它们的细胞构筑、纤维联系和功能各不相同
(一)中央后区
中央后区(postcentral area)发生较晚。于胚胎6个月时才开始分化,出生后才分化完全。此区的边界在灵长类和人则很清楚。它包括中央后回和中央旁小叶后部,相当于3、1、2、43区以及3区和4区间的过渡地带。其中3区和4区间的过渡地带位于中央沟底,其前方为4区,后方为3区,有人称此区为3a区。在细胞构筑上,3区属于颗粒型皮质,含有散在的中、小型锥体细胞,与纹皮质很相似。3区的前部位于中央沟的后壁,称颗粒亚区;3区的后部位于中央沟上缘之后唇,称简单亚区,乃3区的主要代表。1区和2区属于额叶型皮质,内、外颗粒细胞层薄、中央后区的传入纤维主要来自丘脑腹后核的特异性纤维(丘脑中央辐射),它们经由内囊后肢主要投射到3区。1区只接受30%的投射纤维,另外还接受一些到3区去的纤维的侧支。2区主要接受到3区和1区去的纤维的侧支。中央后回与4、6、5、7等区之间有联络纤维,与对侧半球的4、3、1、2、5、7区等处有连合纤维。中央后回的传出纤维是锥体系和锥体外系的重要来源。锥体系约40%的纤维来自中央后回,这些纤维下行止于薄束核、楔束核、三叉神经感觉诸核和孤束核等,构成反馈,从而影响(兴奋或抑制)这些核的上行冲动;躯体感觉区是皮质纹体纤维(锥体外系)的主要起点之一。此外,躯体感觉区还可发出纤维至同侧丘脑腹后核,它们之间的联系是往返的,且有局部定位;皮质网状纤维也源于此区,止于脑干的网状结构。中央后区是浅、深躯体感觉中枢,与躯体各部有明确的定位关系。
1.第Ⅰ躯体感觉区(first somatosensory area,Sml)第Ⅰ躯体感觉区位于中央后回(3、1、2区)并延伸到中央旁小叶后部(图3-2-4)。本区司理躯体的浅、深部感觉。它包括:①指和趾被动运动时的运动感觉和运动方向的感觉(位置觉);②对身体受刺激的部位能精确定位;③对身体上间隔很小的两点,同时施以刺激时,可以辨别出是两个点(两点辨别觉);④感受三维空间(立体)的触觉能力,即能辨别实体的能力;⑤闭眼可以辨别他人在其手、面、躯体和肢体上写的字或画的符号(图形觉);⑥区别物体的轻重(重量觉);⑦辨别物体的软、硬;⑧同时刺激躯体左、右侧,能同时感受到双侧受到刺激。这种感觉既有空间的定位觉,又有时间的定位。第Ⅰ躯体感觉区也接受痛觉、温度觉和轻触觉,不过它们可能在丘脑平面已达到意识阶段,故破坏中央后回时,这些感觉仍然保留。
刺激清醒患者的中央后回,患者主观感觉对侧身体麻木或有触电感觉,有时感到身体某处在运动,但实际上并未运动,极少数可产生痛、温的感觉;口区的感觉是同侧的;咽、喉、会阴等区的感觉可以是双侧的。研究表明3区主要与皮肤浅感觉有关;2区主要与关节的深感觉有关;而1区与浅、深感觉均有关系。
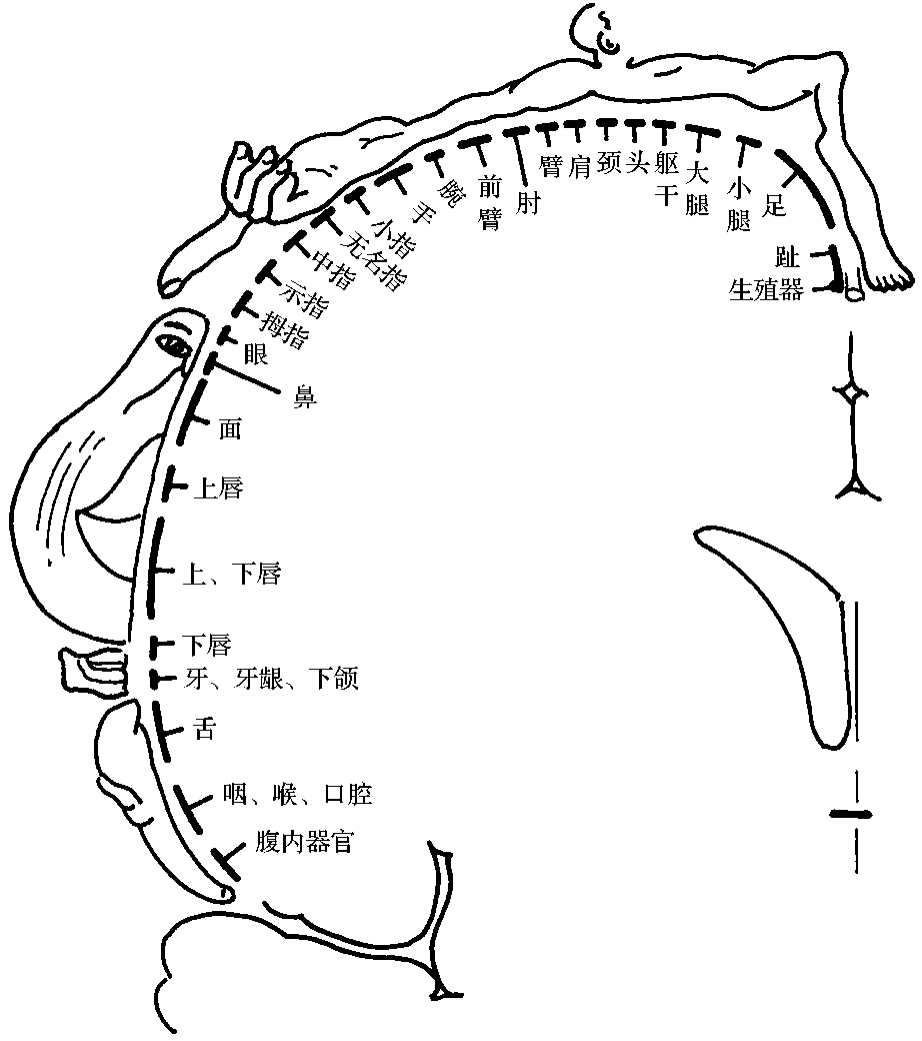
图3-2-4 第一躯体感觉区功能定位示意图
人脑能产生感觉的刺激点,约75%在中央后回,25%在中央前回。刺激周围感受器,中央前回也有电变化,丘脑有直接终于运动区的纤维,切除中央后回,再刺激中央前回,仍可产生感觉,这都说明运动区的感觉运动性及其独立性。但临床上切除中央前回,并不产生任何感觉缺失症状。
刺激中央前、后回,所产生的感觉反应不同。例如使眼产生感觉的刺激点,几乎全在中央前回,使上、下唇产生感觉的刺激点,几乎全在中央后回。位置感觉和运动感觉只见于对侧半球,人脑是否左、右半球都有辨别性触觉代表区,尚未明确。
第Ⅰ躯体感觉区与躯体各部的感觉有精确的对应关系,从半球内侧面开始,沿中央沟自中央旁小叶至大脑外侧沟方向,顺序刺激中央后回和中央前回,患者自足至口、喉依次产生感觉。躯干、四肢是倒置的,与对侧相对应;头面部是正置的,面和舌可能在两侧皮质都有代表区。颈部第8皮节区与面区紧邻。外生殖器的代表区可能在中央旁小叶凹入扣带沟的上唇处,或在半球上缘附近的中央后回上。代表区的大小与该部的感觉灵敏度有关,例如手、面、口的代表区特别大。这是因为感觉灵敏部位有大量的感受器,周围神经分布的密度大,与皮质联系的神经元数量多造成的。
切除全部中央后回,或中央后回严重受损,并不引起瘫痪,但来自骨骼肌、肌腱、关节的位置觉和运动觉以及体表的两点辨别觉均消失,尤以手、足为甚。因此患者不能辨别被动运动的方向和程度,不能分辨物体的轻重、粗细、软硬和温度,不能正确指出触及体表那一点的位置。结果,患者只凭触摸,不能识别他手中的物体,称为实体感缺失(astereognosis)。上述现象只出现于身体对侧。
2.第Ⅱ躯体感觉区(second somatosensory area,SmⅡ)人类SmⅡ位于中央后回最下部并延伸到中央前回。在猴,此区大部深入大脑外侧沟的上唇,邻接岛叶皮质。刺激患者的SmⅡ所得结果与SmⅠ相似,即双侧手、足、上臂有麻木感或瘫痪感。人类此区内的面、舌、口和喉的代表区尚未确定。在猫和猴,身体各部在SmⅡ的定位排列是面部在前,向后依次为上肢、躯干和下肢,双侧都有,但以对侧为主。
此区也与轻触觉以及震颤觉有关。刺激环层小体,SmⅡ所得电位比SmⅠ为高。SmⅡ接受丘脑枕核的传入纤维。传出纤维除投射到丘脑外,也投射到脊髓后柱。此外,与SmⅠ、4区、补充运动区等处有联络纤维;与对侧SmⅡ、SmⅠ间有连合纤维。
附:第三躯体感觉区(thirdsmatosensory area,SmⅢ)
即附加运动区。其感觉功能知之甚少。此区接受丘脑皮质纤维,其传出投射有些纤维不属运动性,但详细情况不太清楚。刺激清醒患者的SmⅢ,据说可引起头部和腹部的一般性感觉,但无确切的局部定位。
3.味觉区(gustatory area)皮质的味觉区尚未完全肯定。一般认为味觉区在中央后回下端,头面部代表区的下方,即顶叶的岛盖部(43区)和邻近的岛叶皮质。切除猴的此处皮质,引起味觉障碍,刺激清醒患者的此区,可产生味觉。在猴的实验中已经查明,味觉经鼓索和舌咽神经传入,由孤束核中继,可能还经臂旁核再次中继,止于丘脑腹后内侧核的小细胞部,最后投射到43区和邻近的岛叶皮质。此区可能有纤维至丘脑腹后核。
(二)顶上区
凡是哺乳动物皆有顶上区(superior parietal area),但灵长类,特别是人,此区最为发达。它位于半球内侧面和背外侧面,有人认为此区仅包括顶上小叶,有人认为它还包括中央旁小叶后半和楔前叶,相当于5区和7区。
本区皮质厚度中等,第Ⅲ层、第Ⅴ层有大的或极大的锥体细胞群是其特征。纤维联系非常复杂,现已查明:丘脑后外侧核与5、7区间有往返纤维,背外侧核与7区间有往返纤维;此外,5区还接受SmⅠ的纤维,5区发出纤维至7区、补充运动区和6区;7区和46区间有往返纤维,此外还发出纤维至顶下小叶和6区等处;顶上区的传出纤维还参与顶枕颞桥束和皮质网状束。与对侧的对称部位有连合纤维。
顶上区的功能较复杂,进化上出现较晚,特别是顶上小叶与对侧肢体精巧的技术性运动有关。它可对来自皮肤、肌腱、关节和内感受器的刺激进行高级的分析综合。例如,它可把中央后回的上肢区和下肢区的简单冲动互相联系起来,辨别肌主动收缩的程度,分辨触觉,区别所感受的压觉,辨别运动的方向和肢体在空间的位置,从而使上、下肢运动得以精确的配合。因此它是与上、下肢精巧活动有关的皮质感觉代表区。若顶上小叶全部损伤,则本体感觉和辨别性触觉,如定位觉、运动方向觉和肢体在空间的位置觉等出现障碍,障碍的程度远比中央后回为重。与中央后回不同者,是此区无局部定位。障碍涉及整个对侧半身,有时同侧也受轻微影响。
三、视区
视区是指与视觉有关的脑区,主要在枕叶皮质。可分为纹状区(17区)、纹旁区(18区)和纹周区(19区)三个区域。过去认为视辐射只终于17区,后来得知外侧膝状体的纤维确实还投射到18区和19区。现在认为枕叶的三个区都与视觉有关,统称视皮质。它们与视网膜的象限有严格的定位关系,三个视皮质区也发出纤维投射到丘脑枕、外侧膝状体和脑干运动核,此外它们和其他皮质区之间也有长的和短的联络纤维,与对侧半球的相应部位有连合纤维。视皮质的面积虽然只占整个大脑皮质总面积的3%,但其神经元数却占半球皮质神经元数的1/10。枕叶功能除视觉外,与眼、头等部位的运动也有一定的关系。
(一)第一视区
第一视区(first visual area)即17区,也称纹状区(striate area)。包括距状沟两侧的皮质区、楔叶和舌回。在人类几乎全部位于枕叶内侧面,猴黄斑的代表区位于枕叶外侧面。17区前界位于胼胝体压部附近,在顶枕沟和距状沟交点前一短距离处,后界通常延展到月状沟。全区面积2 600 mm2,与视网膜面积相比大致为7∶1。皮质厚度较薄,平均1.5 mm。皮质类型属颗粒型,全区结构比较一致,不再分亚型,星状细胞密集,数量大大超过锥状细胞。此区细胞构筑横向分层极其明显,各层细胞可有不同的功能,垂直方向的柱状结构也极明显,同一细胞柱功能相同。
由浅入深可分为:第Ⅰ层,(分子层),仅含少量小型神经元。第Ⅱ层,(外颗粒层),细胞小而密集,胞体多为短锥体形。第Ⅲ层,(外锥体细胞层),细胞数量较多,形状多为锥体形,由浅而深体积逐渐加大。第Ⅳ层,(内颗粒层),很厚,又可分为三个亚层,即Ⅳa、Ⅳb和Ⅳc。Ⅳa也称Gennari上层,细胞为典型的颗粒细胞,较密集。Ⅳb也称Gennari线,或称Baillarger外带,含有大量的视辐射和联络纤维的终末。它们与皮质表面平行,称自有髓纤维,无论是新鲜标本或染色切片均可看到前后纵向的条纹。17区的边界和延展范围虽有个体差异,但当此区过渡到18区时,此种纤维即突然停止,从而构成17区的典型特征,纹状区或纹皮质即由此而得名。Ⅳb的细胞成分相对稀少,但却含有纹皮质中最大的星状细胞,称为Meynert细胞,形成Ⅳb的另一特征。这种细胞通常较为分散,因此有人称为纹皮质的孤立细胞(solitary cells)或巨细胞,它们是锥体细胞的变型,直径30 um,很少出现在Ⅳa和Ⅳc。Ⅳc也称Gennari下层,细胞数量最为丰富。第Ⅴ层,(内锥体细胞层),厚度较薄,细胞体积中等或较大,形态类似Meynert细胞,孤立存在或成群出现,最大者直径可达60 um。第Ⅵ层,(多形细胞层),细胞小而不规则,密集。17区皮质的柱状结构极为明显。每个柱的宽度为25~50 um,相当于1~2个细胞的宽度。每个柱有自己的方位轴,相邻细胞柱的方位轴相差约10°。(www.daowen.com)
从视网膜到枕叶皮质的视觉传导途径主要有两条。一条经外侧膝状体到17区,另一条经上丘和丘脑枕止于18区和19区。膝距束(geniculocalcarine tract)由来自外侧膝状体的纤维组成,此束离开外侧膝状体,经内囊的豆状核后部形成视辐射(optic radiation),大部分止于17区,少部分止于18区和19区。止于17区的部分,主要止于第Ⅳ层,只有少量纤维止于第Ⅰ层。止于18区的部分,终于第Ⅳ层,止于19区的部分,终于第Ⅳ、Ⅴ和Ⅵ层。
视辐射的纤维并非全部都以最短距离达到纹状区。背侧纤维基本上是向后直行,达纹状区。而腹侧纤维,首先在颞叶中行向前下,达侧脑室下角的前部,再返折后行,形成一个纤维袢,此袢名Meyer袢。而后紧靠侧脑室后角外侧壁后行,形成该处的外矢状层,最后达纹状区。视辐射越靠腹侧的纤维,路径越长,最腹侧的纤维所形成的袢,可远达海马旁回、钩。
视辐射的纤维排列有定位。来自外侧膝状体内侧半代表视网膜上象限的纤维,经视辐射的背侧部,投射到距状沟的上唇;来自外侧膝状体外侧半代表视网膜下象限的纤维,经视辐射的腹侧部,投射到距状沟的下唇。来自外侧膝状体中央部代表黄斑区的纤维,占据视辐射中间的大部分,向后终于距状沟后1/3部的上唇和下唇,即17区的后1/3部。此处向前依次为视网膜周围部的代表区。
在实验动物上发现,刺激纹状区的某一点,可在特定的视野区出现简单的视像,例如闪光。刺激视网膜,可在纹状区的单个细胞得到反应。根据突触学说的研究和变性实验的结果得知,投射纤维的冲动传到柱状结构的中间神经元,通过兴奋和抑制的交互作用,经锥状细胞传到皮质下神经元,或传到其他皮质区。每一柱状单位并非完全孤立,它借锥状细胞的返回侧支和分子层的切线纤维又和其他柱状单位相联系。
来自两眼视网膜相称点的纤维止于外侧膝状体同一细胞柱的不同层次,至于相称点冲动的相遇,则发生在纹状皮质中。纹皮质细胞柱在功能与结构上存在着优势现象。纹皮质细胞柱以9个为一组,只接受一个眼的刺激,而邻组细胞柱则接受另一个眼的刺激,如此相间排列。这样,以9个细胞柱为一组相间排列的眼柱群,叫优势眼柱群(ocular dominance)。每个柱的第Ⅳ层只接受一个眼的刺激,而其他层则接受来自双眼的刺激,但以一个眼为主。
外侧膝状体每层只与一个眼(同侧或对侧)的感受野相联系,双眼的相互作用主要在视皮质进行。
上丘是眼球运动的皮质下中枢,在整合各眼外肌运动中起重要作用。它除了直接接受来自视网膜的定位投射、来自视区和额眼区的定位投射(皮质顶盖纤维)以外,还更为广泛的接受来自额、顶、颞叶的纤维;其传出纤维组成顶盖脊髓束下行,参与眼球外肌的运动与头肌运动的协调。部分纤维还上行进入丘脑枕、背内侧核、外侧膝状体、顶盖前区等。丘脑枕直接接受来自视网膜、上丘、顶盖前区、视皮质的纤维,还发出纤维与顶、枕、颞叶联络区有广泛的往返联系。
视皮质除接受外侧膝状体、上丘、丘脑枕的投射纤维以外,17区发出短的联络纤维到18区; 18区发出联络纤维到17区、19区和20区。18区下部的纤维终于19区的上部;18区上部的纤维终于19区的下部。刺激18区可以引起调节反射。19区与额眼区有往返联系,这种联系是眼球借助其随意运动调节视力的结构基础。18区和19区与37区有往返联系,19区与24区有往返联系。视区通过联络纤维和连合纤维和同侧及对侧皮质有联系(两侧17区之间的连合纤维可能很少)。因此,远离视区的皮质损伤,可能影响视觉和眼球的运动。
视区的传出纤维加入皮质顶盖束,主要终于同侧上丘。其定位关系,在短尾猴是:19区的上部纤维至上丘的嘴内侧半;下部纤维至上丘的尾外侧半;17区、18区的上部纤维至上丘的尾外侧半,下部纤维至上丘的嘴内侧半。此外,皮质顶盖纤维还可终于外侧膝状体、顶盖前区和丘脑枕等处。始自视区的皮质被盖纤维还可终于动眼神经核、中介核、后连合核、展神经核和展旁核。展旁核可能是双眼偏斜运动的中枢,右展旁核使双眼偏向右侧,左侧展旁核使双眼偏向左侧。19区的投射纤维有的终于延髓网状结构的抑制区。在视觉系统中,17区的功能是识别物体的立体结构,即形状,不同部分的明暗等。切除人类两侧的17区,视觉丧失,一侧损伤,引起双眼对侧偏盲,均不能恢复。而在某些低等哺乳动物,当第一视区切除后,仍保留大部分视力,它们大概是通过视束至上丘,并由此再至丘脑枕和18区、19区的联系来完成的。切除猴的两侧枕叶,视觉几乎完全丧失,但尚保留粗略的明暗感。若切除猩猩的两侧枕叶,就无任何视觉,且不能恢复。可见在进化中,人类感觉机能上移至大脑皮质,视觉系统达到了极点。但由于视辐射和17区所占面积甚广,在临床上即使是一侧损伤(事实上往往也是不完全的),所以引起的偏盲也多是不完全的(象限性偏盲)。
(二)第二视区和第三视区
1.第二视区(second visual area) 即18区,因位于17区周围,又称为纹旁区(Peristriate area)。皮质较17区厚,属颗粒型。
2.第三视区(third visual area) 即19区,因该区围绕着17区和18区,故又称为纹周区(peristriate area),此区位于枕叶最前部,故也叫枕前区(preoccipital area)。19区前邻7、39、37区。皮质厚度在视皮质中最厚,平均2.5 mm,属顶叶型皮质。
18区和19区内星状细胞不甚显著,锥状细胞更少。在Weigert染色法的切片上观察,一般结构与其他皮质相似,但树突分布野内的纤维和中央后回一样,比较集中。切除18区和19区皮质后,观察外侧膝状体的逆行性变性,其全部六层均可出现柱状细胞群变性,此种现象只有在损坏皮质的全层时才能出现。现已证明18区和19区也有柱状单位,其特点是反应更复杂的刺激,几乎全接受两眼视网膜的冲动。
第一、二视区对感知、整合视觉信息是重要的,常被称为视联络区。动物实验中,光刺激视网膜,在此二区可见到诱发电位。损伤人类18区和19区,患者很难识别物体的形状、大小及其意义。损伤优势半球的此二区,还可引起阅读困难,患者难于理解所见到的文字的意义。
人类除17、18、19区与视觉有肯定的关系而外,颞叶的某些部位也与视觉及其相关活动有某些关系。临床观察得知额叶癫痫、肿瘤或刺激这些部位,这些患者可有幻视。而且,切除该部位后,有关幻觉的记忆依然存在。临床研究发现右侧颞叶(非优势半球)与快速视觉辨认有关,其前部与视觉引起的复杂行为有关,其后部(颞上沟处)与18区有联络纤维,也与视觉有某些关系。枕叶皮质除视觉的特化而外,刺激枕叶的一些部位还可引起眼球向同侧偏斜、头部转动、面肌运动,有时上肢也有反应。说明视区的特化虽然相当高级,但也是感觉运动性的,有人称之为枕眼区(occipital eye field)。
四、听区
听区是指与听觉有关的脑区主要是颞叶皮质。颞叶皮质包括新皮质、古皮质、旧皮质和过渡型皮质。海马旁回位于底面内侧部,属旧皮质和过渡型皮质。海马和齿状回原位于底面的内侧部,由于发育缓慢而被卷入侧脑室下角的底壁,属于古皮质,见于嗅脑和边缘叶。此处仅涉及颞叶的新皮质部分。
颞叶新皮质在种系发生上出现较晚,在人类获得了高度的发展,占颞叶的大部分,居外侧面和底面的外侧部。一般认为它与听、语言、知觉和记忆等有关。
听区(acoustic area)在人脑有第一听区和第二听区,在猫还有第三听区,位于前二者的后方。
(一)第一听区
相当于41区,位于颞横前回的中部和颞横后回的一部分。细胞构筑属于变异的颗粒型皮质,比视区和躯体感觉区的皮质厚,约3 mm。所有各层都含小细胞。Ⅰ层细胞甚少;Ⅱ、Ⅲ层多由小锥形细胞组成,Ⅳ层的细胞密度更大一些;Ⅴ层较厚,颗粒细胞密集成团,略排列成柱;Ⅴ层在Nissl染色切片上染色浅,小的和中等的细胞排列成柱;Ⅵ层由许多多形细胞和锥形细胞组成,但不成柱。
(二)第二听区
位于AI区的外侧,相当于42区及其邻接的22区,占颞横回的其余部分以及邻接的额上回。42区属顶叶型皮质,且第Ⅳ层柱状结构明显,第Ⅲ层内有许多大锥形细胞。22区细胞有排列成柱的倾向。
(三)听区的纤维联系
41区是主要的听接受区;42区是主要的听联络区;与41区、42区相邻接的22区的一部分也是听联络区。听区的传入投射主要来自内侧膝状体的小细胞部发出的听辐射(auditory radiation)。听辐射也叫膝颞纤维(geniculotemporal fibers),它们主要终于41区,其中来自内侧膝状体小细胞和大细胞部的少量纤维终于42区。42区又借联络纤维与41区,特别是22区相联系。
听皮质对音频有定位。听辐射背侧部纤维主要传导来自对侧耳蜗底的高音冲动,止于41区的后内侧部;听辐射腹侧部纤维主要传导来自对侧耳蜗顶的低音冲动,止于41区的前外侧部。
听皮质的音频配布可能反映在细胞的柱状结构上,每一细胞柱对一种特定的频率最敏感。但也有人发现同一细胞柱又可对非常不同的频率发生反应。
听区的联络纤维主要是41区发出纤维至42区;41区、42区发出纤维至22区。此外,41区、42区还可发出纤维至额眼区(8区)、视联络区(18区、19区)、额叶和顶叶的某些区域(6、44、43、1区)、21区和37区等。22区接受的联络纤维最多,而它又与额叶的44区、45区、9区、10区和8区,与额叶和顶叶的岛盖部,与顶叶的39区,与颞叶的21区、38区、37区,与枕叶的18区、19区有广泛的联系。由此可知听皮质与语言皮质、躯体感觉、视皮质等有密切关系。
听区的连合纤维经胼胝体和前连合既可与对称部位相联系又可与不对称部位相联系。听区的传出纤维中的下行投射与上行通路相伴行。它们大部分起自第一听区,经双侧下丘的中央周核、延髓的橄榄周核中继,再经第Ⅷ脑神经至双侧考蒂器的毛细胞,但以对侧为主,对上行冲动起加工(加强信号和抑制噪音)作用,是一种反馈控制。
此外,听皮质也发出纤维至上丘,经顶盖桥束至背外侧桥核,中继后至小脑蚓的视听区。此通路是诸多复杂的听皮质—小脑通路之一。听皮质的下行投射还可至脑神经运动核,构成听觉引起的多种反射的结构基础。其中有些纤维可能影响镫骨肌和鼓膜张肌的收缩,与提高声音的频率有关。
颞叶新皮质的功能虽然研究很多,但尚未完全了解。听觉是颞叶比较肯定的功能。第一听区能感受简单的声音,如铃声、哨声等,但比较复杂的声音则由联络区整合。作为对特种声音的语言的理解,也在联络区内进行,特别是在优势半球一侧的联络区。联络区的后部是Wernicke区的一部分,与语言关系至为密切(见顶叶的后说话区)。
由于内侧膝状体的纤维来自双侧,故单侧内侧膝状体和41区、42区损伤,只能引起患侧听力的轻度下降,甚至颞叶广泛损伤,对听力影响也不大。但单侧损伤后,不易判断声音的来源,特别是声音的远近。有人报道双侧听区破坏可致全聋,但也有听觉不受影响的相反报导。颞叶损伤主要表现在语言功能上,特别是优势半球。双侧22区损伤,可以导致严重的听觉性失语,这可能是因为听觉冲动传至双侧皮质的缘故。此外,左侧额叶后部损伤,常出现遗忘性失语,患者不能叫出一个熟悉的人或物的名称,即命名不能(anomia)。
刺激优势半球的颞上回和邻近颞中回的22区以及颞叶后部延入角回、缘上回的部分时,可出现语言停止现象,与刺激中央前、后回、额上回制止说话区的效应相似。
五、语言区
在动物界,人类的语言功能进化到极其复杂和高等的程度。语言的构成在脑区是一个综合功能的表现。它需要对视觉、听觉接受的信号进行处理,更需要把处理后的信号用语言形式表达出来。与语言有关的脑区涉及额叶、顶叶和颞叶等结构。在功能上主要分为说话区和说话抑制区。
(一)说话区
第一语言区(first speech area)通常只有优势半球(左侧)才有说话代表区。它们是前、上和后说话区。前和上说话区位于额叶。后说话区位于顶叶和颞叶。
1.前说话区(anterior speech area)即运动性语言区(the motor speech area),是第Ⅰ躯体运动区向下后部延伸的部分,相当于44区和45区的一部分。1861年,法解剖学家兼外科医生Broca报道2例左侧大脑半球额下回部损伤,伴有失语症。这一现象被后人的临床观察和实验所证实,故称此区为Broca回(区)[Broca's gyrus(area)]。损伤此区,一般会出现运动性失语症(motor aphasia)。运动性失语症只是拼音不清楚,与发音有关的骨骼肌并未瘫痪。刺激相邻的4区,还可引起肌收缩或简单的发音。这种失语症,有的人不能恢复,有的人可以逐渐恢复正常,这说明优势现象不是绝对的。有的人没有优势半球现象,有的人此区位于右侧半球(图3-2-5)。
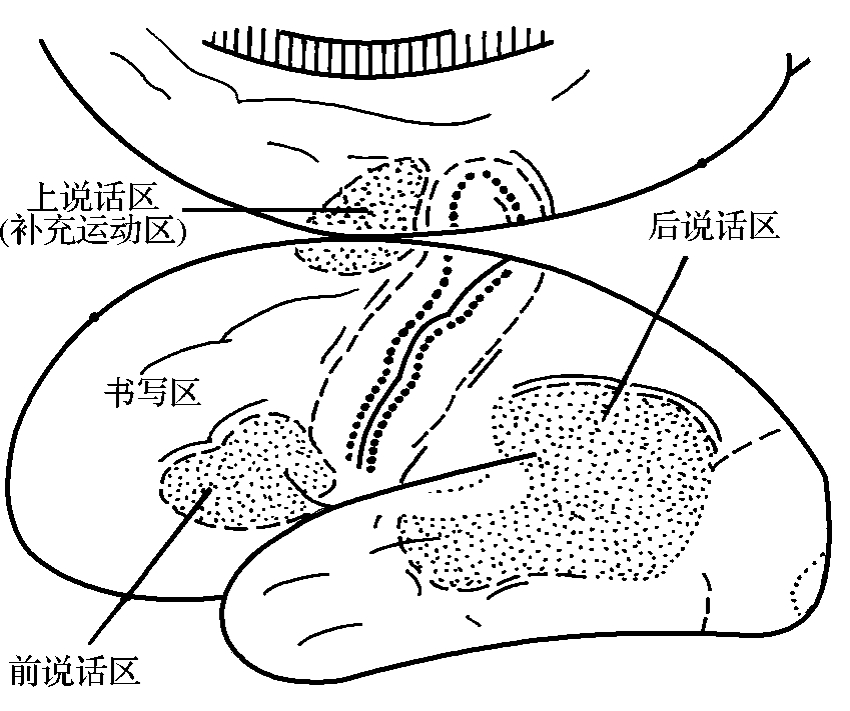
图3-2-5 左侧半球语言区示意图
2.上说话区(superior speech area) 亦称补充说话区(supplementary speech area),位于半球内侧面的额内侧回。损伤此区,失语症仅维持数周即可恢复正常。
3.后说话区(posterior speech area) 或称Wernlcke区,又称第二语言区(second speech area)。第二语言区仅占顶叶的一小部分,大部分位于颞叶。顶叶部分大致相当于39区和40区,颞叶部分位于颞上回后部(22区)和颞中回后部(37区)。第二语言区和5区、7区(与躯体感觉有关),41区、42区(与听觉有关),18区、19区(与视觉有关)间有丰富的联络纤维;和对侧有连合纤维;和丘脑背侧核、后外侧核间有往返纤维;此外还参与皮质脊髓束和皮质被盖束。后者影响脑干运动核,特别是眼肌运动核。第二语言区是人脑最重要的语言中枢,其重要性远大于Broca区,破坏此区的后果比破坏Broca区严重,故一般认为在手术时除非万不得已不要破坏此区。此区损伤后的失语症叫感觉性失语症(sensory aphasia)。颞上回后部是听觉性语言中枢。此区受损,患者能听到别人讲话的声音,但听不懂别人谈话的意思。结果是答非所问,语无伦次,这种情况叫字聋。此时,其他语言功能,如看字、写字、说话均正常。角回是视觉性语言中枢。此区受损,患者虽然能看见物象或文字,但不能对人、物命名或理解字的意义,即不能阅读,这种情况叫命名性失语(anomic aphasia)或失读症(alexia)。此时,患者其他语言功能,如说话、听话、写字均正常。实际上很难见到纯运动性或纯感觉性失语症,最常见到的是混合性的。
4.与语言有关的脑区 顶下区除与语言有重要关系外,优势半球的顶下小叶还与书写、计算、辨别左右方向、辨别自己手指的顺序、阅读和技巧运动有关,非优势半球的顶下小叶还与感知对侧半身的存在、方位感、地形关系感等有关。
上述可见顶上区和顶下区是重要的皮质区,它们与邻近的感觉皮质间有着密切的联系,一般感觉和特殊感觉均在此进行复合和整合,它们具有高度分析综合的能力。临床上损伤或切除此区,除发生严重的感觉性失语症(优势半球损伤)、对侧上、下肢精巧运动障碍而外,还可出现种种混杂现象,如肌无力,说话反常和空间定位觉消失。特别是实体觉的缺失与损伤顶叶有关,即在没有视觉的帮助下,不能用手检查物体的形状、大小和重量。
5.辅助说话区 每侧半球各有两个电刺激可以引起发音的区域。它们是中央前、后回发音区和额上回发音区。
(1)中央前、后回发音区(central vocalization) 通常称为Rolando发音区,位于中央前、后回,与唇、下颌、舌代表区重叠。刺激中央前回引起发音的次数,四倍于中央后回。有些人在发音的同时伴有双唇、面肌、下颌、舌和咽的运动,或伴有口腔和舌的感觉。切除此区仅产生构音障碍(dysarthria),不导致失语症。
(2)额上回发音区(superior frontal vocalization) 位于半球内侧面的额内侧回,中央沟向前4cm处,与补充运动区部分重叠。电刺激时仅少数人引起发音。
(二)说话抑制区
每侧半球各有两个区域,电刺激时,正在进行的语言可以突然停止。它们是中央前、后回制止说话区和额上回制止说话区(图3-2-6)。
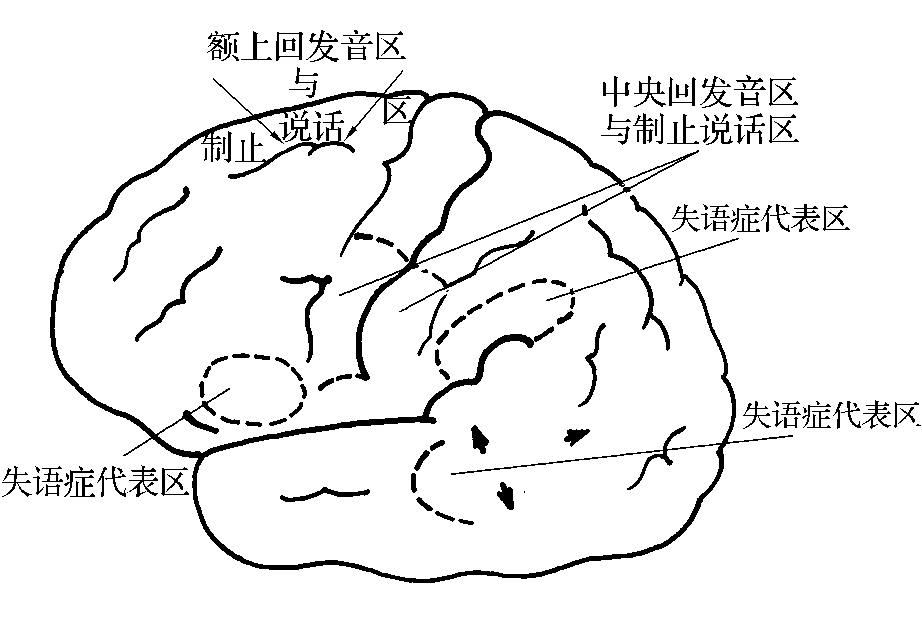
图3-2-6 左侧半球皮质的发音区与制止语言区
1.中央前、后回制止说话区(central speech arrest area) 位于中央前、后回的喉、面代表区之间,与中央前、后回发音区相重叠。刺激中央前回引起说话中止的次数,也四倍于中央后回。
2.额上回制止说话区(superior frontal speech arrest area) 位于半球内侧面,与额上回发音区相当。
(三)书写区
书写区(writing area)位于优势半球的额中回后部(8区)。此区损伤的患者,不能以书写的方式表达自己的意见,但手部的其他运动功能仍然存在,阅读和说话的功能也不受影响,这种情况称为失写症(agraphia)。
人类的上、前、后说话区,或说、写、看、听四个语言中枢,善用右手(右利)的人位于左侧半球;大部分善用左手(左利)的人,也位于左半球,只有一部分左利的人位于右半球。与语言有关的半球通常称为优势半球。优势半球的此区受损,有时对侧半球可代偿其功能。在一般情况下,非优势半球的这些区域可能是未活动的语言区。
通过PET检查图像证实,上述三个语言区或四个语言中枢不是孤立的,它们的功能除与联络纤维、连合纤维有关而外,更有赖于和丘脑之间的联系。仔细切除Broca区和Wernike区的灰质层,而不损伤其深侧的白质,结果不产生失语症。但左侧丘脑枕出血或丘脑肿瘤而无皮质损伤的患者,却有严重的失语症。从剥制纤维的脑标本上就可看到Broca区与丘脑背内侧核、中央中核有纤维联系,补充说话区与丘脑枕有纤维联系,Wernike区与丘脑枕有纤维联系。现在的观点是丘脑枕也参与说话中枢。即说话的功能与皮质说话区、丘脑以及它们之间的纤维联系三者有关(图3-2-7)。
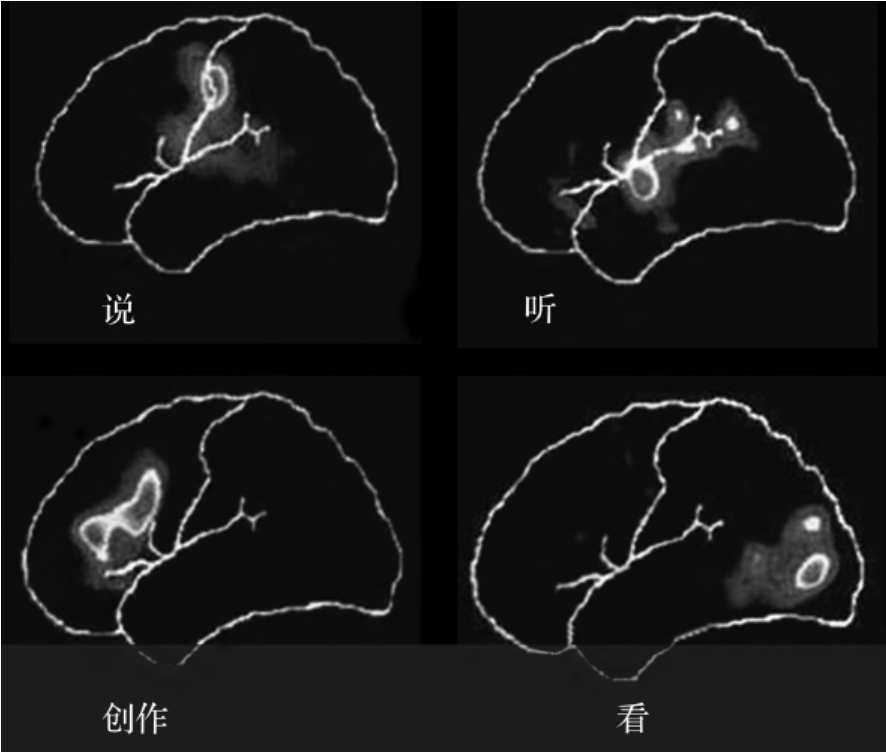
图3-2-7 语言中枢的PET扫描图像
Damasio等近来提出大脑皮质内有三套处理语言信息的装置。即概念系统、语言系统和介导系统。概念系统位于两侧额叶的前区和中间区,其功能是摄取各种感觉形成概念,此区受损,则不能形成事物的概念。语言系统位于优势半球,即Broca区和Wernike区,其功能是说话。此区受损,造成失语症。介导系统的功能是摄取概念,诱导产生语言,或者是接受语言,刺激概念系统产生新的概念,它们位于优势半球的枕颞轴上。一般性概念(如普通名词)、颜色概念位于颞叶后区,特殊性概念(如专用名词)位于颞极。介导系统受损,患者彻底不能运用所有的名词,不能对熟人和颜色命名,但仍保持形成概念的能力。以上至少说明除三个语言区或四个语言中枢之外,确实还有其他的皮质区参与语言的加工。应该指出,所谓“优势半球”是指语言和文字、符号等方面有所特化的一侧半球,一般指左侧半球。而“非优势半球”(一般指右侧半球),现在已经知道它们在空间感觉、美术、音乐等方面有所特化,例如认识人的面貌、凭借外形认识物体,识别音乐的主旋律等。因此实际上两个半球各有所长,功能上是互补的,无所谓“优势”和“非优势”[7]。
六、内脏脑
内脏脑(visceral brain)是指以边缘系统为代表的通过下丘脑影响一系列的自主神经活动的神经结构而言。边缘系统由边缘叶(扣带回、海马旁回和钩、海马、齿状回)与边缘叶皮质结构相似的皮质(额叶眶回后部、岛叶前部、和颞极)以及皮质下结构(隔核、杏仁核族、下丘脑、上丘脑、丘脑前核以及中脑被盖内侧区等)构成。边缘叶在发生上比较古老,在功能上和纤维联系上十分密切。它不仅与嗅觉有关,更与内脏活动、情绪行为活动和记忆等密切相关(见边缘系统)。
(李振平)
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。





