厌氧发酵又称沼气化、沼气发酵、甲烷发酵等,是指有机物(如禽兽粪便、稻秆、有机废水、能源作物等)在厌氧细菌的作用下进行代谢产生以甲烷为主的可燃气体沼气的过程。沼气是一种优质、清洁、高效的气体燃料,经净化和提纯,甲烷含量可提高至95%,成为与天然气各项指标都相同的生物天然气。与好氧生物处理相比,沼气厌氧发酵处理具有较多优势:可以有效降解高浓度的固体废弃物;处理工艺设计和建设成本低,基建面积小,能源消耗少;反应处于厌氧条件下,可以有效抑制病原菌等的滋生及减少有毒有害臭气的释放;产生的沼气可以作为清洁能源,沼渣沼液可以回收再利用。目前,沼气生产以“产业沼气”为中心,向规模化、集约化的方向发展。产业沼气以大中型沼气工程为生产形式,以大量的沼气或生物天然气为产品,由管道或气罐瓶经济地输送和装运,实现大规模产业化应用。
一、沼气发酵原理
沼气是一种混合气体,主要成分是甲烷,其次有二氧化碳、硫化氢、氮气和其他一些成分。沼气的组成中,可燃成分包括甲烷、硫化氢、一氧化碳和重烃等气体;不可燃成分包括二氧化碳、氮气和氨等气体。在沼气成分中甲烷含量为55%~70%、二氧化碳含量为28%~44%、硫化氢平均含量为0.034%。1g COD在厌氧条件下完全降解可以生成0.25g甲烷,相当于标准状态下的甲烷气体体积为0.35L。沼气厌氧发酵可利用的底物种类多样,不同类型底物发酵产沼气量及沼气的主要成分如表12-10所示。
表12-10 不同底物沼气产量及沼气主要成分
沼气发酵的过程是微生物在特定的厌氧条件下,将有机物质进行分解的过程,其中一部分碳素转化成CH4和CO2。在这个转化过程中,被分解的有机碳化合物的能量大部分转化并储存在CH4中,仅小部分有机碳化合物氧化为CO2。关于沼气发酵有两阶段理论、三阶段理论和四阶段理论之说。20世纪30年代,沼气发酵被概括为产酸阶段和产甲烷阶段,即两阶段理论;1970年,Bryantlzgl等在两阶段理论的基础上,突出了产氢产乙酸菌的作用;同时,Zeikuslao等提出四类群理论,突出同型产乙酸菌的功能。复杂有机物的沼气发酵过程以及各阶段的碳源转化率如图12-11所示(其中Ⅰ、Ⅱ、Ⅲ为三阶段理论,Ⅰ、Ⅱ、Ⅲ、Ⅳ为四阶段理论)。
目前,普遍接受并认同的是三阶段理论,即沼气发酵过程主要分为三个阶段:水解发酵阶段、产氢产乙酸阶段和产甲烷阶段。
(1)水解发酵阶段厌氧发酵的原料几乎都是不溶性有机大分子纤维素、蛋白质和脂肪等,只有通过水解酶将其分解为小分子,而后才能被产氢产乙酸菌和产甲烷细菌利用,最终合成为甲烷。多种微生物可以产生胞外酶,如水解酶有脂肪酶、蛋白酶和纤维素酶等。它们的功能是将复杂的大分子水解为可被微生物同化的小分子。首先各种不溶性原料被发酵性细菌所分泌的胞外酶水解为简单的可溶性的有机物;然后由厌氧和兼性厌氧的水解性细菌或发酵性细菌将多糖水解成单糖或二糖,将蛋白质转化成肽和氨基酸,将脂肪酸转变为甘油和脂肪酸,并进而形成乙酸、丙酸、丁酸等脂肪酸与醇类以及一定量的CO2、H2等。研究表明水解发酵阶段较为缓慢,特别是纤维素类物质的水解。该阶段是沼气发酵过程的限速步骤。
图12-11 复杂有机物的沼气发酵过程及各阶段的碳源转化率
(2)产氢产乙酸阶段四阶段理论中将产氢产乙酸阶段分为产氢产乙酸过程及同型产乙酸过程。(a)在产氢产乙酸过程中,产氢产乙酸细菌(H2-production acetogens)可将产酸发酵阶段产生的挥发性有机酸和醇转化为乙酸、H2/CO2,主要微生物类群有脱硫弧菌属(Desulfovibio)和梭菌属(Clostridium spp.)等。(b)在同型产乙酸过程中,同型产乙酸细菌(homo-acetogens)可利用挥发性水解发酵阶段的产物有机酸、H2和CO2为底物还原为乙酸,主要微生物类群有乙酸梭菌(C.aceticum)、伍迪乙酸杆菌(Acetobacterium woodii)等。在这个阶段主要的产物是乙酸,约占70%以上。乙酸为产生大量甲烷奠定了物质基础,有70%的甲烷都是在发酵过程中由乙酸形成的。因此这一阶段是复杂的有机物转换成沼气的先决条件。在产氢产乙酸阶段,产甲烷菌可以利用产氢产乙酸菌代谢过程中产生的氢,使丙酸、丁酸和乙醇的氢分压降低;而同型产乙酸菌可通过还原反应使系统氢分压降低,从而使反应可自发进行,确保产酸发酵过程的正常进行。
(3)产甲烷阶段此阶段的反应由严格厌氧的产甲烷菌群(methanogens或methane-producingbacteria)来完成。这类微生物只能利用一碳化合物、乙酸和氢气形成甲烷。在其形成过程中,约有30%的甲烷是来自H2的氧化和CO2的还原;另外70%的甲烷则来自乙酸盐的转化。由于产氨细菌的大量活动,使氨态氮浓度增高,氧化还原势降低,为甲烷菌生活提供适宜的厌氧环境。再由严格厌氧的产甲烷菌群将第二阶段分解出来的乙酸等简单有机物分解成甲烷。产甲烷菌是自然界碳素循环中厌氧生物链的最后一个成员,对自然界物质循环起着重要的作用。
在沼气发酵过程中,上述三个阶段并不是完全孤立和分开的,而是相互连接、交替进行的。它们之间保持动态平衡。
二、沼气发酵微生物
在沼气发酵过程中,不同的微生物菌群通过各自的生理代谢途径起着不同的物质转化作用,且相互协同共同完成有机废弃物的厌氧消化过程。根据沼气发酵微生物的代谢功能的不同,主要划分为非产甲烷菌(non-methanogens)和产甲烷菌(methanogens)两大类群。根据其呼吸作用不同,可将非产甲烷菌分为厌氧菌、兼性厌氧菌和好氧菌三大类群,其中厌氧菌是产酸发酵阶段的主要微生物类群;根据代谢产物的不同,非产甲烷菌还可被分为发酵细菌、产氢产乙酸细菌和同型产乙酸细菌三大类群。产甲烷菌属于严格厌氧菌,在厌氧发酵中利用乙酸、甲醇或甲胺等一碳化合物及H2和CO2生成甲烷。产甲烷菌通常被划分为三大类群:氢营养型产甲烷菌、乙酸营养型产甲烷菌和甲基营养型产甲烷菌,不同菌群在厌氧消化的不同时期构成优势功能菌群。沼气发酵过程实际是这些微生物区系相互偶联反应的过程,在各种化学偶联中非产甲烷菌和产甲烷菌相互依赖,又互为制约关系。主要关系表现为:不产甲烷菌主要为甲烷菌群提供生长繁殖的基础物质和厌氧条件,以及消除部分有毒物质,同时产甲烷菌群消耗掉不产甲烷菌群的代谢产物,解除反馈抑制,创造热力学上的有利条件,并共同维持适宜的pH。
1.非产甲烷菌
非产甲烷细菌主要有三类:发酵性细菌、产氢产乙酸菌和同型产乙酸菌。这三类微生物共同完成水解发酵阶段和产酸产氢阶段。
(1)发酵性细菌 该类菌群的主要功能是在厌氧条件下将多种复杂有机物水解为可溶性物质,并将可溶性有机物发酵生成乙酸、丙酸、丁酸、氢和二氧化碳等物质。发酵细菌大多数为专性厌氧菌,也有大量兼性好氧及兼性厌氧菌。由于厌氧发酵原料的多样性与复杂性,发酵性细菌也是一个复杂的混合菌群。按其代谢功能,发酵细菌可分为纤维素分解菌、半纤维素分解菌、淀粉分解菌、蛋白质分解菌和脂肪分解菌。目前已探知的发酵细菌有几百种,主要包括梭菌属(Clostridium)、拟杆菌属(Bacteroides)、丁酸弧菌属(Butyrivibrio)、真杆菌属(Eubacterium)和双歧杆菌属(Bifidobacterium)等专性厌氧细菌;兼性厌氧菌包括链球菌属(Streptococcus)和一些肠道细菌等。
厌氧消化过程中,发酵性细菌将大分子不溶性有机物水解成小分子的水溶性有机物。水解过程是在菌体胞外分泌的胞外水解酶作用下完成的。水解产物被微生物菌体吸收后,经过细胞内复杂的酶催化系统作用,将一部分作为能源使用的有机物转化为代谢产物,分泌到细胞外的水溶液中,被参与下一阶段生化反应的微生物吸收利用(有机酸、醇、酮等)。
(2)产氢产乙酸菌 发酵性细菌将复杂有机物分解发酵所产生的有机酸和醇类,除乙酸、甲酸和甲醇外均不能被产甲烷菌所利用,必须由产氢产乙酸菌将其分解。产氢产乙酸过程主要是微生物将两个碳以上的有机物(除乙酸外)和醇转化为乙酸、氢气和二氧化碳等,并产生新的细胞物质的过程。主要反应过程如下:
研究表明,产氢产乙酸菌是一类严格厌氧的真细菌,并且与产甲烷菌是互营细菌,即其生长和代谢完全依赖于产甲烷菌等其他微生物。由于氢气的反馈抑制作用,除乳酸降解外,丁酸、丙酸等有机酸均不能降解,必须依靠产甲烷菌将其代谢产物如H2转移利用。在营养生态位上位于产酸发酵菌群和产甲烷菌群之间,在功能生态位上起到承上启下的重要作用,能将产酸发酵菌群代谢产生的丙酸、丁酸等挥发性有机酸(VFA)和乙醇等进一步降解转化为乙酸和CO2/H2,为后续的产甲烷菌群提供可以直接利用的底物。产氢产乙酸菌生长和代谢依赖于代谢产物氢或甲酸的消除,因而必须与产甲烷菌和硫酸盐还原细菌之类的耗氢细菌组成共培养体系才能维持生长。产氢产乙酸菌主要有脱硫弧菌(Desulfovibio desulfuricans)、普通脱硫弧菌(Desulfovibio vulgaris)、沃尔夫互营单胞菌(Syntrophononas wolfei)、沃林互营杆菌(Syntrophobacter wolinii)等。
(3)同型产乙酸菌也称为耗氢产乙酸菌,是一类既能自养生活又能异养生活的混合营养型细菌,既能利用H2和CO2生成乙酸,也能代谢糖类产生乙酸。同型产乙酸细菌可以产生乙酸,为产甲烷菌提供了形成甲烷的基质;在代谢过程中消耗H2而降低H2分压,对产H2的发酵细菌有利,使厌氧消化系统中保持较低的H2分压,有利于沼气发酵正常进行。由葡萄糖转化为甲烷的化学式如下:
C6H12O6+2H2O → 2CH3COOH+2CO2+4H2
4H2+CO2→ 2H2O+CH4
CH3COOH → CO2+CH4(https://www.daowen.com)
理论上由乙酸形成的甲烷占微生物产甲烷总量的67%。因为1分子葡萄糖产生2分子乙酸和4分子氢气,产生1分子甲烷需要4分子氢气或者1分子乙酸,所以在厌氧降解碳水化合物形成甲烷的过程中,通过氢气产生的甲烷最多,占总甲烷的33%。常见的同型产乙酸菌主要有伍迪乙酸杆菌(Acetobacterium woodii)、威林格乙酸杆菌(Acetobacterium wieringae)、乙酸梭菌(Clostridium aceticum)、嗜热自养梭菌(Clostridium thermoautotrophicum)等。
2.产甲烷菌
产甲烷菌是一类形态多样,具有特殊细胞成分,可以代谢H2和CO及少数几种简单有机物生成甲烷的严格厌氧古生菌。它们不能利用氧气作为电子受体,只有少数产甲烷古生菌在微量氧气中短时间存活,是厌氧消化过程中的最后一组成员。产甲烷菌在严格厌氧的条件下将前三类非产甲烷细菌代谢的产物,在没有外源受氢体的情况下,把乙酸和H2/CO2转化为CH4、CO2和水,使有机物在厌氧条件下的分解作用得以顺利完成。
产甲烷菌广泛分布于自然界,在污泥、瘤胃、人和动物肠道、昆虫肠道、地热水泉、深海火山口、海洋及淡水沉积物、水田和沼泽等厌氧环境中都有产甲烷菌的存在。产甲烷菌属于广古生菌门(Euryarchaeota),分布在7个目中:甲烷杆菌目(Methanobacteriales)、甲烷微菌目(Methanomicrobiales)、甲烷八叠球菌目(Methanosarcinales)、甲烷球菌目(Methanococcales)、甲烷火球菌目(Methanopyrales)、甲烷胞菌目(Methanocellales)和Methanomassiliicoccales。目前分离鉴定的产甲烷菌已有200多种。有6个目的产甲烷菌基因组信息被阐明(表12-11)。根据已有的数据,产甲烷菌基因组大小一般介于(1.6~5.7)×106bp,G+C含量为30%~65%。
表12-11 部分产甲烷菌及其基因组信息
根据所利用的底物不同,可将产甲烷菌分为三大类群。(a)氢营养型:甲烷杆菌科(Methanobacteriaceae)、甲烷球菌科(Methanococcaceae)、甲烷微菌科(Methanomicorbiaceae)、甲烷盘菌科(Methanoplanaceae)和甲烷嗜热菌科(Methanothermaceae),除代谢H2和CO2外,大约有一半的种还可以代谢甲酸。氢营养型产甲烷菌利用氢的效率很高,表现出对H2的极大亲和力。甲烷八叠球菌常被视作乙酸营养型产甲烷菌的代表,实际上它们是混合营养型的,大多数菌株以H2为主要能源,对乙酸的亲和力相当低。(b)乙酸营养型:该类型产甲烷菌的真正代表是索氏甲烷丝状菌,它只利用乙酸作为CH4产生的底物,并能有效地利用乙酸,可将所吸收乙酸的98%~99%的甲基转化成甲烷,因此是厌氧颗粒污泥和淡水沉积物等生境中最重要的产甲烷菌。(c)甲基营养型:一些甲基类化合物(如甲醇、三甲基胺、二甲酰硫化物)以及其他的一些醇(如异丙醇、异丁醇、乙醇、环戊醇等)也是某些产甲烷菌的底物。但是专一性的甲基营养菌数量不多,如甲基拟甲烷球菌(Methanococcoides methylutens)和丁达尔甲烷叶菌(Methanolobus tindarius)等,它们只利用甲醇和甲胺生成甲烷。目前研究结果表明,以甲基化合物为原料合成的甲烷还不足10%。甲烷合成过程中氢营养型产甲烷途径所需自由能最高,其次是甲基营养型产甲烷途径,所需自由能最少的是乙酸营养型产甲烷途径。作为有机物厌氧发酵最后阶段的核心功能微生物类群,产甲烷菌在沼气厌氧发酵中发挥至关重要的作用。在不同的环境中,不同的产甲烷菌参与分解有机物,分解对苯二甲酸盐(TA)的产甲烷菌主要是乙酸营养型的甲烷鬃毛菌属(Methanosaeta)和还原二氧化碳营养型的甲烷微菌目。甲烷冷泉碳酸盐岩和淤泥中主要的产甲烷菌是甲烷鬃毛菌属。分解链烃的产甲烷菌主要是氢营养型,高温条件下的产甲烷菌主要是氢营养型和甲基营养型。
产甲烷菌种类多样,形态不同,但它们具有共同的生理特性与功能:①生长要求严格厌氧环境,对氧气和氧化剂非常敏感,在有氧气的条件下就不能生存。经测定,环境中的氧化还原电位高于-0.33V时,产甲烷菌则不能生长。②只能利用少数简单的化合物作为营养,仅限于利用H2/CO2、甲醇、甲酰、乙酸及甲基胺等为底物生成甲烷。所有产甲烷菌均以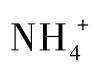 作为主要氮素来源,生长均需要微量的镍、钴、钼和铁,这些微量元素是产甲烷菌特有辅酶的重要成分。③产甲烷菌适合生长于中性偏碱性条件。在厌氧消化器中,当pH低于5.5时,沼气发酵就会完全停止。④产甲烷菌生长非常缓慢。产甲烷菌代谢的底物是乙酸、甲酸、H2/CO2等结构简单含能量少的物质,生活于严格的厌氧条件下,其代谢产物CH4含有很高的能量,所以生长繁殖缓慢。
作为主要氮素来源,生长均需要微量的镍、钴、钼和铁,这些微量元素是产甲烷菌特有辅酶的重要成分。③产甲烷菌适合生长于中性偏碱性条件。在厌氧消化器中,当pH低于5.5时,沼气发酵就会完全停止。④产甲烷菌生长非常缓慢。产甲烷菌代谢的底物是乙酸、甲酸、H2/CO2等结构简单含能量少的物质,生活于严格的厌氧条件下,其代谢产物CH4含有很高的能量,所以生长繁殖缓慢。
三、产甲烷菌形成甲烷的途径
1.独特的辅酶系统
产甲烷菌中含有原核生物类群中许多独特的辅酶。这些辅酶在产甲烷作用的生物化学中起着重要作用,许多辅酶比其他原核生物中许多普通辅酶如NAD +或FMN的含量高得多。根据其功能的不同,这些辅酶可以分为两类,一类是C1的携带者,主要包括甲烷呋喃(MFR)、四氢甲烷蝶呤(H4MPT)、四氢八叠甲烷蝶呤(H4SPT)和辅酶M(HS-CoM);另一类是电子载体,主要包括铁氧化还原蛋白(Fd)、辅酶B(HSCoB)、辅酶F430、细胞色素、FAD、甲烷吩嗪(MP)和辅酶F420。
(1)产甲烷作用中作为C 1载体的辅酶甲烷呋喃(MFR)是一种低分子质量的辅酶,它参与甲烷形成作用的第一步反应,在产甲烷作用的最初CO2被还原成甲基水平的同时与呋喃的氨基端结合,并且在这个过程的最后转移给第二个辅酶。H4MPT和H4SPT类似,均为四氢叶酸的衍生物,用于甲酰、亚甲基和甲基水平上C1的转移,其转移方式与四氢叶酸类似。在产甲烷途径中大多数的还原步骤中,甲烷蝶呤携带着C1单位由甲酰水平到甲基水平。辅酶M(CoM)是一种能参与产甲烷菌甲基转移反应的酶,具有异常的含硫量和很强的酸性。由它组成的甲基辅酶M还原酶是一种仅存在于多种产甲烷菌细胞中,催化二氧化碳还原为甲烷的最后一步反应的酶分子。辅酶M是甲基基团的携带者,通过F430甲基还原酶复合物在甲烷形成作用的最后一步将甲基还原成甲烷。
(2)氧化还原反应中的辅酶辅酶F430是一种黄色可溶性含有镍的四吡咯,并且类似CoM,辅酶F430作为甲基还原系统的一部分在甲烷形成的末端步骤中起关键作用。HS-CoB(7-磷酸巯基庚基苏氨酸)包含一个巯基和磷酸基团,其中巯基为其活性部位,这个辅助因子被包括在通过甲基还原系统催化甲烷作用的最后一步,作为电子供体参与甲基还原系统。MP是一种仅存在于甲烷八叠球菌目中的一种辅酶,其功能与其他细菌呼吸链中的醌类相似。辅酶F420是一种去氮黄素衍生物,在波长为420nm的紫外光下发出蓝绿色荧光,可以利用这一特点初步鉴定是否存在产甲烷菌细胞。辅酶F420的电势较低,既可作为氢化酶、甲酸脱氢酶和一氧化碳脱氢酶的电子受体,又可作为NADP +还原酶的电子供体。
产甲烷菌在电子传递磷酸化作用中,缺乏细胞色素和醌(泛醌和甲基萘醌),更没有植物体中的叶绿素和动物体中的血红素,但产甲烷菌体内却有一种特殊的电子传递体,就是辅酶F420、F430和甲烷蝶呤等。这些辅酶是产甲烷菌特有的电子传递系统,在生理活动中起着电子传递的作用。
2.产甲烷菌的生化代谢途径
产甲烷菌的生化代谢途径如图12-12所示,其中从CO2出发,垂直向下至CH4的代谢途径代表由H2和CO2形成甲烷途径,从左侧CH3COOH出发,向右至CH3—H4M(S)PT,再向下至CH4的路径为乙酸产甲烷途径,从右侧甲基化合物出发,至CH4—S—CoM,再向下至CH4的路径代表甲基裂解途径。在甲烷生成代谢途径中,以CH3-S-CoM产生甲烷过程是三种代谢途径所共有的,它是由含镍的甲基辅酶M还原酶(Mcr)催化,以HSCoB为直接的电子供体还原CH3-S-CoM产生甲烷和CoB-S-SCoM的过程。其中CoB-S-S-CoB被视为产甲烷过程的末端电子受体,它被异质二硫化物还原酶(Hdr)还原,重新释放出HS-CoB和HS-CoM,其反应方程式为:CoB-S-S-CoM+2e -+2H +→CoB-SH+CoMSH。
图12-12 产甲烷古生菌的生化代谢途径
(1)由H2和CO2形成甲烷途径CO2首先被甲酰甲烷呋喃脱氢酶还原为甲酰基,并连接在C 1载体甲烷呋喃(MFR)上形成CHO-MFR。这是一个吸能反应,需要跨膜离子或质子梯度来驱动。—CHO被转移给另一个C 1载体四氢甲烷蝶呤(H4MPT),再依次被还原为次甲基(≡CH)、亚甲基(═CH2)和甲基(—CH3)。催化这些反应需要辅酶F420作为电子载体。在甲基四氢甲烷蝶呤:CoM甲基转移酶(Mtr)的作用下,甲基被转移到第3个C1载体巯基辅酶M(HS-CoM)上,这个跨膜蛋白与Na +跨膜传输偶联,从而在还原CO2的过程中推动形成跨膜的Na +梯度。在辅酶M与辅酶B的作用下甲基基团最终脱落形成CH4。最后一步是甲基辅酶M还原酶(Mcr)将其转化为甲烷。
(2)乙酸产甲烷途径自然界中约2/3的生物产甲烷来自于乙酸途径,然而仅Methanosarcina和Methanosaeta能够利用乙酸途径产生甲烷。在乙酸发酵产甲烷途径中,Methanosarcina和Methanothrix激活1个乙酸产生乙酰辅酶A,分别需要消耗1个和2个ATP。Methanosarcina通过乙酸激酶(AK)和磷酸转乙酰酶(PTA)生成乙酰辅酶A。Methanothrix利用乙酰辅酶A合成酶(ACS)和焦磷酸化酶(PPase)活化乙酸。其中Methanosarcina和Methanothrix激活1个乙酸分别需要消耗1个和2个ATP。它们再通过CO脱氢酶/乙酰辅酶A脱羧酶复合体(Codh/Acs),氧化乙酸上的羧基为CO2,转移乙酸中的甲基到辅因子四氢八叠甲烷蝶呤(H4SPT)上,其中,Codh/Acs是乙酸途径中的关键酶,用于剪切乙酰辅酶A上的C—C和C—S键。在将甲基基团转移到H4SPT上形成CH3-H4 SPT的同时,利用Fdox作为电子受体氧化CO产生CO2和Fdred。甲基被还原为甲烷的过程与CO2还原途径一致。
(3)甲基裂解途径首先甲基由甲基转移酶复合体激活和转运,后者包括2个甲基转移酶。甲基转移酶1(MT1)将底物上的甲基转移到MT1上与类咕啉蛋白结合,甲基转移酶2(MT2)再转移甲基与HS-CoM连接产生CH3-S-CoM。在H2存在的条件下,CH3-S-CoM利用H2作为电子供体产生甲烷;在没有H2的条件下,一部分CH3-CoM经过CO2还原逆途径被氧化产生CO2、F420H2和H2,另一部分CH3-S-CoM利用F420H2和H2作为电子供体产CH4。
四、沼气发酵过程中“超复杂”物化反应过程
由于厌氧发酵涉及一系列复杂的微生物代谢反应,而反应体系又存在固、液、气三种物态,除了微生物生理生化反应外,还涉及复杂的物化反应过程,包括酸碱解离平衡、固液溶液与沉淀平衡、气相传质平衡及氨的利用等。液相中的物化过程主要涉及氢离子、氢氧根离子、有机酸根、铵离子以及碳酸氢根等之间的离解平衡,其反应速率非常快,可直接用稳态的平衡方程式来表达。在厌氧反应过程中,会有少量的硫化物或碳酸根存在;同时进水或反应器中也会同时存在一定量的各种金属离子,如Ca2 +、Fe2 +、Zn2 +等。这些阴阳离子共存于厌氧反应器中,会生成沉淀;同时随着反应器内混合液的pH、温度等的变化,又会存在这些沉淀物的溶解与沉淀之间的平衡。厌氧消化过程所产生的沼气中主要含有CH4、H2、CO2等三种气体组分。由于氨极易溶于水,反应过程中产生的氨全部存在于液相中,在气相中不存在氨气组分;同时,由于进水中还可能含有少量的硫酸盐,硫酸盐还原作用的进行可能会导致气体组分中还会含有微量的H2S;对CH4、H2、CO2等气体来说,传质阻力主要在液相。由于大部分CH4和CO2逸出,氨以亚硝酸铵、碳酸氢铵的形式留在污泥中,它们可中和第一阶段产生的酸,为产甲烷菌创造了生存所需的弱碱性环境。氨可被产甲烷菌用作氮源。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。





