自然界中约118种化学元素以单质或化合物的形式构成了地球上形形色色的物质,其中碳、氢、氧、氮、磷、硫、铁等20种化学元素是构成自然界中生物体生命活动物质基础的必需元素。生物地球化学循环(biogeochemical cycle)是指生物圈中的化学元素经过各种物理、化学及生物化学作用在生命体与其周围环境之间的迁移和循环,尤其是生物化学作用。生物本身在生物地球化学循环中起着重要的推动作用,其中构成生物体内糖类、脂质、蛋白质和核酸等有机物大分子的主要化学元素(C、H、O、N、P、S)在自然界中循环较快。
微生物在整个生态系统中的物质循环、能量流动及信息传递过程中发挥着重要作用,其代谢活动也相应地改变着其周围生境的物理和化学特性,进而影响生态系统的功能。微生物在生态系统中可以扮演多种角色并行使多种功能。①微生物是生态系统的初级生产者。一些自养微生物能固定太阳能、无机物的化学能,并将简单的无机物合成为有机物,进而使得固定下来的能源在食物链和食物网中进行单向流动,逐级递减;而微生物合成的有机物与其生境之间进行物质循环。②微生物是有机物的主要分解者。微生物在生态系统中的最大价值在于其强大的分解功能,它们能将植物、动物及微生物残体中的复杂有机物最终分解为简单无机物以供初级生产者利用。地球上90%以上有机物的无机质化都是由微生物完成的。③微生物是物质和能量的储存者。土壤和水体等环境中含有大量的微生物并储存着大量的物质和能量。④微生物是生态系统中的信息接受者和信息源。微生物可以从环境中接收信息,主要表现为趋光性和趋化性,同时微生物也可以向生物体和环境发出信息,如饥饿状态下的黏细菌(Myxococcus xanthus)通过信号分子传递信息,引起细菌出芽及聚集,进而实现黏细菌之间的合作共赢。
一、碳循环
从全球的观点来讲,碳元素以多种形式(如CO2、 、HCO3、CaCO3及各种有机物等)存在,并在自然界碳储库中以不同的速率进行循环。自然界中的碳储库主要有大气、陆地、海洋、沉积物、岩石(主要指石灰岩)及化石燃料等(图7-8)。岩石是地球上最大的碳储库(储量约1.2×1017t),比海洋及大气中的碳含量分别高4个和6个数量级,但其中碳元素的循环速率十分缓慢。大气虽然是自然界中最小的碳储库,但它作为不同碳储库之间碳元素迁移的主要载体,循环最快。海洋通过吸收和储存大气中的碳元素影响着全球碳元素循环,其中人类活动导致的排放中的30%~50%被海洋吸收。但是,随着石油和煤等化石燃料燃烧量的日益增加,大气中CO2含量持续升高,导致了全球性的“温室效应”和碳元素循环失衡,并由此引起了气候变暖、海平面上升、土地沙漠化等一系列全球性环境问题,进而威胁到全人类的正常生产和生活环境。目前大气中的CO2浓度已达到400mg/L,比工业时代之前的大气CO2浓度增加了50%。社会生物地球化学模型(socio-biogeochemical models)预测21世纪中叶大气中CO2浓度将达到550mg/L,到2100年大气中CO2浓度将超过700mg/L。
、HCO3、CaCO3及各种有机物等)存在,并在自然界碳储库中以不同的速率进行循环。自然界中的碳储库主要有大气、陆地、海洋、沉积物、岩石(主要指石灰岩)及化石燃料等(图7-8)。岩石是地球上最大的碳储库(储量约1.2×1017t),比海洋及大气中的碳含量分别高4个和6个数量级,但其中碳元素的循环速率十分缓慢。大气虽然是自然界中最小的碳储库,但它作为不同碳储库之间碳元素迁移的主要载体,循环最快。海洋通过吸收和储存大气中的碳元素影响着全球碳元素循环,其中人类活动导致的排放中的30%~50%被海洋吸收。但是,随着石油和煤等化石燃料燃烧量的日益增加,大气中CO2含量持续升高,导致了全球性的“温室效应”和碳元素循环失衡,并由此引起了气候变暖、海平面上升、土地沙漠化等一系列全球性环境问题,进而威胁到全人类的正常生产和生活环境。目前大气中的CO2浓度已达到400mg/L,比工业时代之前的大气CO2浓度增加了50%。社会生物地球化学模型(socio-biogeochemical models)预测21世纪中叶大气中CO2浓度将达到550mg/L,到2100年大气中CO2浓度将超过700mg/L。
从生物体的观点来看,碳元素是构成所有生物体中有机物质的最主要元素,约占有机物干重的50%,是整个生态系统中碳循环的中心,被植物、藻类、蓝细菌及化能自养微生物利用进行光合作用和化能合成作用,形成有机化合物。其中,植物性有机化合物被动物摄入后合成动物性有机化合物。植物和动物通过呼吸作用将其部分有机化合物分解形成的CO2及厌氧微生物和好氧微生物将动植物残体中的有机化合物彻底分解产生的CO2重新释放到大气中,进而形成完整的碳循环(图7-9)。微生物参与了地球生态系统中碳循环的各个环节,其作用可以概括为作为生产者参与固定大气CO2合成有机物的过程,及作为分解者参与将生境中的有机物最终分解为CO2的过程。
图7-8 自然界中碳元素的分布与循环
图7-9 碳元素的氧化还原循环
1.微生物在CO2固定过程中的作用
碳元素的生物固定是指生物将自然界中的无机碳(CO2和 )转变为稳定有机物的过程,主要分为还原性(自养式)碳固定和非还原性(异养式)碳固定,两者之间的一个重要区别是在碳元素固定过程中是否进行了碳元素的还原。在还原性碳固定过程中,无机碳可以作为唯一碳源经过一系列反应最终被还原成糖类等生命物质,同时伴随着无机碳结合受体再生,如植物、藻类及自养微生物利用光能或化学能将无机碳最终还原成为生命物质(尤其是糖类)的过程。在非还原性碳固定过程中,无机碳仅被固定在相应的有机物受体上而非单独地形成有机物,如草酰乙酸盐途径作为异养生物三羧酸循环途径的回补途径,该过程中CO2在丙酮酸羧化酶的作用下形成草酰乙酸。
)转变为稳定有机物的过程,主要分为还原性(自养式)碳固定和非还原性(异养式)碳固定,两者之间的一个重要区别是在碳元素固定过程中是否进行了碳元素的还原。在还原性碳固定过程中,无机碳可以作为唯一碳源经过一系列反应最终被还原成糖类等生命物质,同时伴随着无机碳结合受体再生,如植物、藻类及自养微生物利用光能或化学能将无机碳最终还原成为生命物质(尤其是糖类)的过程。在非还原性碳固定过程中,无机碳仅被固定在相应的有机物受体上而非单独地形成有机物,如草酰乙酸盐途径作为异养生物三羧酸循环途径的回补途径,该过程中CO2在丙酮酸羧化酶的作用下形成草酰乙酸。
目前在微生物中已发现的自养式碳固定途径有六条:卡尔文循环(Calvin-Benson cycle)途径、厌氧乙酰辅酶A途径(anaerobic acyty-CoA pathway)、逆向三羧酸循环途径(reversed tricarboxylic acid cycle)、3-羟基丙酸途径(3-hydroxyproprionate pathway)、3-羟基丙酸/4-羟基丁酸循环途径(3-hydroxypropionate/4-hydroxybutyrate cycle)及二羧酸/4-羟基丁酸循环途径(dicarboxylate/4-hydroxybutyrate cycle),其中后两条途径发现最晚,分别由Berg和Huber两位科学家在2007年和2008年报道。每种途径中所涉及的代表性微生物见表7-3。碳固定过程中的能量来源于生物的氧化磷酸化、底物水平磷酸化、环式光合磷酸化及非环式光合磷酸化等过程。
表7-3 碳固定途径中所涉及的代表性微生物
续表
(1)卡尔文循环途径 卡尔文循环又称Calvin-Benson-Bassham(CBB)循环及还原性戊糖磷酸循环(reductive pentose phosphate cycle),是自然界自养生物中最为普遍存在的CO2固定途径,包括真核生物中的绿色植物和藻类及原核生物中的蓝细菌、许多好氧或兼性好氧的变形菌门(含α-变形菌纲、β-变形菌纲及γ-变形菌纲)、硫细菌、铁细菌及消化细菌等光能和化能自养微生物(表7-3)。
在卡尔文循环中,核酮糖-1,5-二磷酸羧化酶/加氧酶(RubisCO或RuBPCase,ribulose 1,5-bisphosphate carboxylase/oxygenase)及核酮糖磷酸激酶(phosphoribulokinase)是两个特征酶,其中RubisCO酶被认为是地球上含量最为丰富的酶。到目前为止,共发现4种结构类型的RubisCO酶,即Ⅰ型、Ⅱ型、Ⅲ型和Ⅳ型,其中RubisCO酶主要为Ⅰ型,存在于所有植物、藻类、蓝细菌及大多数化能自养微生物中。该型RubisCO酶由两类明显不同的亚基,即大亚基(50~55ku)和小亚基(12~18ku)组成。Ⅱ型RubisCO酶仅在少量的细菌中发现。该型酶仅由大亚基组成,其中Rhodospirillum rubrum仅含有Ⅱ型RubisCO酶。也有一些微生物既含有Ⅰ型RubisCO酶,也含有Ⅱ型RubisCO酶,如Rhodobatter capsulatus、Hydrogenovibrio marinus和Rhodobacter sphoeroides等。此外,能进行羧化反应的酶为Ⅰ型、Ⅱ型和Ⅲ型RubisCO酶,而Ⅳ型RubisCO酶参与其他反应。
卡尔文循环过程可分为三个阶段(图7-10),即CO2固定期、1,3-二磷酸甘油酸还原期及CO2固定受体再生期。整个循环过程中的起始化合物是作为CO2固定受体的核酮糖-1,5-二磷酸(RuBp)。
第一个阶段是CO2固定期,又称羧化反应阶段,即在RuBisCO酶的作用下,3分子的CO2结合在3分子CO2固定受体(核酮糖-1,5-二磷酸,RuBp)上面,然后经过不稳定中间代谢物过渡态最终形成了6分子的3-磷酸甘油酸(PGA)(图7-11)。
第二个阶段是1,3-二磷酸甘油酸还原期,是被固定CO2的还原反应阶段。在该阶段,第一阶段形成的3-磷酸甘油酸上面的羧基立即进行还原反应,即在3-磷酸甘油激酶和3-磷酸甘油醛脱氢酶的作用下,3-磷酸甘油酸被还原成3-磷酸甘油醛(图7-12)。
图7-10 卡尔文循环及有机物生成
虚线框部分表示来源于卡尔文循环产生的3-磷酸甘油醛可以经一系列复杂的生化反应合成有机物质
图7-11 卡尔文循环CO2固定反应
图7-12 卡尔文循环的1,3-二磷酸甘油酸还原期
如图7-10所示,6分子的1,3-二磷酸甘油酸被还原成6分子的3-磷酸甘油醛,其中5分子的3-磷酸甘油醛用于CO2固定受体再生,而1分子的3-磷酸甘油醛用于后续有机物的合成。因此,在卡尔文循环过程中,尽管CO2固定的直接产物是3-磷酸甘油醛,但它是糖类物质合成的起点,同时也是其他有机化合物合成的基础,包括有机酸、脂肪及氨基酸等。此外,在3-磷酸甘油醛经一系列生化反应转化为糖的过程中,己糖尤其是葡萄糖是主要合成的糖类。因此,常把葡萄糖当作CO2固定的主要终产物,其净反应式为:
6CO2+12NAD(P)H+12H ++18ATP→C6H12O6+6H2O+12NAD(P)++18ADP+Pi
第三个阶段是CO2固定受体再生期,即第二阶段产生的部分3-磷酸甘油醛先经缩合及糖分子重排等一系列复杂反应形成5-磷酸核酮糖,然后5-磷酸核酮糖在磷酸核酮糖激酶的催化作用下转变为核酮糖1,5-二磷酸,可以重新用于CO2固定,保证卡尔文循环持续进行(图7-13)。
图7-13 卡尔文循环CO2固定受体再生期
(2)厌氧乙酰辅酶A途径 厌氧乙酰辅酶A途径(anaerobic acetyl-CoA pathway),又称Wood/Ljungdahl(WL)途径或还原性乙酰辅酶A途径(reductive acetyl-CoA pathway)。该条CO2固定途径是在专性厌氧的醋酸菌中发现并被阐明的,且主要存在于厚壁菌门及广古生菌门专性厌氧细菌和古生菌[包括醋酸菌,如热乙酸梭菌(Clostridiumthermoaceticum)、醋酸梭菌(C.aceticum)及伍氏醋酸梭菌(Acetobacterium woodii)]、绝大多数的自养型硫酸盐还原菌,如自养脱硫杆菌(Desulfobacterium autotrophicum)及古生菌中甲烷菌,如嗜热自养甲烷杆菌(Methanobacterium thermoautotrophicum)。
尽管厌氧乙酰辅酶A途径非常简单,即将两分子的CO2合成为乙酸,但是根据每消耗1个ATP所能固定CO2的能力来讲,WL途径相比其他五条途径是最高效的CO2固定途径。该途径每形成1分子的乙酰辅酶A,需要两分子的CO2、1分子的ATP及4分子的NAD(P)H,即:
2CO2+1ATP+4NAD(P)H+H ++HS-CoA→乙酰-CoA+1ADP+4NAD(P)+
该途径不同于其他五条碳固定途径,而是采用非循环方式对CO2进行固定,且可以分为两个支路,即甲基支路和羰基支路(图7-14)。整个途径中的关键酶为一氧化碳脱氢酶(CO dehydrogenase,CODH)/乙酰辅酶A合成酶(acetyl-CoA synthase,ACS)的双功能金属酶,即该酶既能发挥CO脱氢酶的作用,又能行使合成乙酰辅酶A的功能。此外,该酶存在于能利用WL途径的所有原核生物中且对氧气敏感。在甲基支路反应中,1分子的CO2被固定在四氢蝶呤(tetrahydropterin,H4PT)辅因子上面,其中在醋酸菌中的辅因子为四氢叶酸(tetrahydrofolate,THF),而在甲烷菌中的辅因子为四氢甲烷蝶呤(tetrahydromethanopterin);CO2被还原力[H](通过F420因子或NADP酶的转移)最终还原成甲基与辅因子的组合(CH3-H4PT)。在羰基支路中,1分子的CO2被一氧化碳脱氢酶还原成羰基并结合在该酶的Ni原子上面,与一氧化碳脱氢酶形成复合体(Ni-CO)。最后,甲基支路反应形成的甲基与羰基支路形成的羰基在乙酰辅酶A合成酶的作用下合成乙酰辅酶A。
图7-14 厌氧乙酰辅酶A途径
在甲基支路中,实线箭头表示醋酸菌将CO2还原成CH3-H4PT的步骤,而虚线箭头代表甲烷菌将CO2还原成CH3-H4PT的步骤
Fd—铁氧化还原蛋白THF—四氢叶酸H4PT—四氢蝶呤(醋酸菌中为四氢叶酸,甲烷菌中为四氢甲烷蝶呤)
MFR—甲基呋喃 F420—辅酶F420
途径中数字代表的酶分别为:1—甲酸脱氢酶 2—甲酰-THF合成酶 3—甲酰-MFR脱氢酶
4—甲酰-MFR/H4PT甲酰转移酶 5—甲酰-THF环水解酶 6—甲酰-H4TPT环水解酶
7—亚甲基-THF脱氢酶 8—亚甲基-H4TPT脱氢酶 9—亚甲基-THF还原酶
10—亚甲基-H4TPT还原酶 11—CO脱氢酶/乙酰-CoA合成酶
乙酰辅酶A是能量和物质代谢途径中的重要物质,在生物体内能量和物质代谢中发挥着枢纽的作用。乙酰辅酶A可以通过一系列复杂的生化反应合成细胞所需的有机物质,例如,乙酰辅酶A与CO2分子在丙酮酸合成酶的作用下合成丙酮酸,丙酮酸通过糖异生途径合成葡萄糖。
此外,甲烷菌和醋酸菌利用WL途径不但能固定CO2,同时也能利用产生的电化学梯度保存能量。在自养生长或发酵来保存能量的过程中,醋酸菌产生乙酸,而甲烷菌通过回收WL途径甲基支路中的CH3-H4PT,然后在辅酶M的作用下将其还原为甲烷。
(3)逆向三羧酸循环途径 逆向三羧酸循环途径,又称反克雷布斯循环(reverse Krebs cycle)或还原性羧酸循环(reductive carboxylic acid cycle)。该途径首先在绿色硫细菌Chlorobium limicola提出,后来发现该途径也存在于其他无氧或微氧条件下的微生物中(表7-3),如绿色硫细菌中的Chlorobium thiosulfatophilum,硫还原细菌中的脱硫杆菌属(Desulfobacter)及古生菌泉古生菌门中的热变形菌属(Thermoproteus)等。
还原性三羧酸循环是氧化性柠檬酸循环的逆向反应。在三羧酸循环过程中1分子乙酰辅酶A被降解成2分子的CO2并产生能量,而在其逆反应中,合成1分子的乙酰辅酶A需要2分子ATP和来源于NADH及NADPH的H +。逆向三羧酸循环途径中应涉及的主要物质有柠檬酸、草酰乙酸、苹果酸、延胡索酸、琥珀酸、琥珀酰辅酶A、α-酮戊二酸及异柠檬酸(图7-15)。本循环起始于作为CO2固定受体的草酰乙酸,每循环一周可羧化固定2分子CO2,最终还原成可供各种生物合成用的乙酰辅酶A。该途径及乙酰辅酶A进一步合成丙酮酸、磷酸烯醇式丙酮酸(PEP)及草酰乙酸所需的酶,如图7-15所示,其中大多数酶与氧化性三羧酸循环途径的酶相同,其中的关键酶有依赖ATP的柠檬酸裂解酶,能将柠檬酸(6C化合物)裂解成草酰乙酸(4C)、乙酰辅酶A、依赖铁氧还原蛋白(Fd)的丙酮酸合成酶和α-酮戊二酸合成酶。乙酰辅酶A在丙酮酸合成酶的作用下结合1分子CO2合成丙酮酸,丙酮酸经生化反应可以进一步形成丙糖和己糖等构成细胞所需要的重要合成原料。
(4)3-羟基丙酸双循环途径3-羟基丙酸双循环途径CO2固定途径首先是由Helge Holo在光合自养型绿色非硫细菌Chloroflexus aurantiacus中发现的,直到后来Georg Fuchs等人彻底阐明了该途径,因此,又称为Fuchs-Holo双循环途径,且是绿色非硫细菌(如Chloroflexus)及一些古生菌(如Sulfolobus和Acidianus)特有的CO2固定途径(表7-3)。起初,乙醛酸(glyoxlate)被认为是Chloroflexus spp.利用该途径固定CO2的主要产物。但乙醛酸并非细胞物质合成的中心前体代谢物,其要转变为细胞中心前体物质需要第二个循环(图7-16),其中3-羟基丙酸为特征性中间代谢物。因此,该循环被定义为3-羟基丙酸双循环。3-羟基丙酸途径与自养生物中其他CO2固定途径的不同特征主要为:是一个双循环偶联的代谢过程,涉及多个步骤及双功能和多功能酶并产生了多种中间代谢产物。
本循环途径涉及的主要化合物有14种,包括乙酰辅酶A、丙二酸单酰辅酶A、3-羟基丙酸、丙酰辅酶A、α-甲基丙二酸单酰辅酶A、琥珀酰辅酶A、苹果酸(malate)、苹果酰辅酶A、乙醛酸、β-甲基苹果酰辅酶A、甲基延胡索酰-C1-辅酶A、甲基延胡索酰-C4-辅酶A、α-甲基苹果酰-CoA及丙酮酸(图7-16)。本途径19个反应步骤需要13种酶,10a、10b及10c是一个三功能酶,即苹果酰-CoA/β-甲基苹果酰-CoA/α-甲基苹果酰-CoA裂解酶(图7-16)。该途径中的一系列反应在有氧条件下均可以进行,涉及的酶在有氧条件下可行使催化功能。
图7-15 绿色硫细菌中的逆向三羧酸循环途径
实线箭头方向表示逆向三羧酸循环途径,本循环起始化合物为草酰乙酸,终产物为乙酰-CoA。除逆向三羧酸循环途径外,乙酰-CoA经同化作用生成丙酮酸、磷酸烯醇式丙酮酸(PEP)及草酰乙酸的途径也整合入该图(虚线箭头方向)
rTCA循环及乙酰-CoA同化过程中所需的酶分别为:1—ATP依赖型柠檬酸裂解酶 2—苹果酸脱氢酶
3—延胡索酸水合酶 4—延胡索酸还原酶 5—琥珀酰-CoA合成酶
6—铁氧化还原蛋白(Fd)依赖型2-酮戊二酸合成酶 7—异柠檬酸脱氢酶 8—乌头酸水合酶
9—Fd依赖型丙酮酸合成酶 10—磷酸烯醇式丙酮酸合成酶 11—PEP羧化酶
在3-羟基丙酸途径中,第一个循环主要用于乙醛酸的合成,起始化合物为乙酰辅酶A,产物为乙醛酸。首先乙酰辅酶A结合溶解形态的CO2(即 )进行羧化反应,合成丙二酸单酰辅酶A。然后丙二酸单酰辅酶A先还原为3-羟基丙酸,并进一步被还原为丙酰辅酶A,其中3-羟基丙酸为第一个循环的特征性中间代谢物。丙酰辅酶A经过羧化反应和分子重排作用进一步形成苹果酰辅酶A,其为该循环的第二个特征性中间代谢物。最终,苹果酰辅酶A在三功能酶(MMC裂解酶)的作用下可以再生为起始化合物乙酰辅酶A,同时形成产物乙醛酸(图7-16)。在第二个循环中,反应起始化合物为乙醛酸和丙酰辅酶A,终产物为丙酮酸和乙酰辅酶A。首先乙醛酸在MMC裂解酶的作用下结合到丙酰辅酶A上面,形成β-甲基苹果酰-CoA,其再经一系列生化反应形成α-甲基苹果酰-CoA,其在MMC裂解酶的作用下生成丙酮酸和乙酰辅酶A,乙酰辅酶A可以重新生成丙酰辅酶A,进而形成了第二个循环(图7-16)。3-羟基丙酸双循环固定CO2途径需要能量高,每生成1分子丙酮酸需要7分子的ATP及3分子碳酸氢根离子。此外,通过糖异生途径丙酮酸还需要额外的3分子ATP形成磷酸丙糖。
)进行羧化反应,合成丙二酸单酰辅酶A。然后丙二酸单酰辅酶A先还原为3-羟基丙酸,并进一步被还原为丙酰辅酶A,其中3-羟基丙酸为第一个循环的特征性中间代谢物。丙酰辅酶A经过羧化反应和分子重排作用进一步形成苹果酰辅酶A,其为该循环的第二个特征性中间代谢物。最终,苹果酰辅酶A在三功能酶(MMC裂解酶)的作用下可以再生为起始化合物乙酰辅酶A,同时形成产物乙醛酸(图7-16)。在第二个循环中,反应起始化合物为乙醛酸和丙酰辅酶A,终产物为丙酮酸和乙酰辅酶A。首先乙醛酸在MMC裂解酶的作用下结合到丙酰辅酶A上面,形成β-甲基苹果酰-CoA,其再经一系列生化反应形成α-甲基苹果酰-CoA,其在MMC裂解酶的作用下生成丙酮酸和乙酰辅酶A,乙酰辅酶A可以重新生成丙酰辅酶A,进而形成了第二个循环(图7-16)。3-羟基丙酸双循环固定CO2途径需要能量高,每生成1分子丙酮酸需要7分子的ATP及3分子碳酸氢根离子。此外,通过糖异生途径丙酮酸还需要额外的3分子ATP形成磷酸丙糖。
图7-16 Chloroflexus(绿弯菌属)固定CO2的羟基丙酸途径
该途径中涉及的酶分别为:1—乙酰-CoA羧化酶 2—丙二酸单酰-CoA还原酶 3—丙酰-CoA合成酶
4—丙酰-CoA羧化酶 5—甲基丙二酸单酰-CoA异构酶 6—甲基丙二酸单酰-CoA变位酶
7—琥珀酰-CoA/苹果酰-CoA转移酶 8—琥珀酸脱氢酶 9—延胡索酸水合酶
10a、10b及 10c—三功能性的酶(MMC裂解酶)(S)—苹果酰-CoA(a)/
β-甲基苹果酰-CoA(b)/(S)α-甲基苹果酰-CoA裂解酶(c)
11—甲基延胡索酰-C1-CoA水合酶 12—甲基延胡索酰-CoA C1-C4 CoA转移酶
13—甲基延胡索酰-C4-CoA水合酶
(5)3-羟基丙酸/4-羟基丁酸循环途径(3-HP/4-HB) 该循环途径发现于泉古生菌门嗜热嗜酸硫化叶菌目(Sulfolobales)中的勤奋金属球菌(Metallosphaera sedula),同时也存在于该目中的其他古生菌,如Stygiolobus azoricus。3-HP/4-HB途径可以分为两个部分,第一部分是从乙酰辅酶A形成琥珀酰辅酶A阶段;第二部分是由琥珀酰辅酶A再生为乙酰辅酶A的过程,其中乙酰辅酶A可以用于多种细胞物质合成的前体。本循环途径中涉及的主要代谢物有15种,包括乙酰辅酶A、丙二酸单酰辅酶A、丙二酸半醛、3-羟基丙酸、3-羟基丙酰辅酶A、丙烯酰辅酶A、丙酰辅酶A、α-甲基丙二酸单酰辅酶A、琥珀酰辅酶A、琥珀酰半醛、4-羟基丁酸、4-羟基丁酰辅酶A、巴豆酰辅酶A、3-羟基丁酰辅酶A及乙酰辅酶A(图7-17)。
3-HP/4-HB循环途径中的第一部分(从乙酰辅酶A到琥珀酰辅酶A)反应顺序与嗜热光全绿丝菌(C.aurantiacus)中3-羟基丙酸双循环途径是一致的(图7-17右侧部分)。但是从丙二酸单酰辅酶A转变为丙酰辅酶A的过程中,两条途径中所涉及的酶存在较大的差异,即3-HP/4-HB途径需要5种酶(图7-17),而3-HP途径仅需要两种酶(图7-16)。在3-HP/4-HB循环途径中的第二部分(从琥珀酰辅酶A再生乙酰辅酶A)中,琥珀酰辅酶A在多种酶的作用下经4-羟基丁酰辅酶A逐渐转变为乙酰乙酰辅酶A,进而裂解为两分子的乙酰辅酶A,这与3-HP双循环途径明显不同;其中4-羟基丁酰辅酶A脱氢酶为该部分中最为重要的酶。该途径存在多种功能酶,如乙酰-CoA/丙酰-CoA羧化酶能催化乙酰辅酶A和丙酰辅酶A的羧化反应。
图7-17 硫化叶菌目(Sulfolobales)3-羟基丙酸/4-羟基丁酸循环途径
本途径涉及的酶:1—乙酰-CoA/丙酰-CoA羧化酶 2—丙二酸单酰辅酶A还原酶
3—丙二酸半醛还原酶 4—3-羟基丙酰辅酶A连接酶 5—3-羟基丙酰辅酶A脱氢酶
6—丙烯酰辅酶A还原酶 7—α-甲基丙二酸单酰辅酶A异构酶 8—α-甲基丙二酸单酰辅酶A变位酶
9—琥珀酰辅酶A还原酶 10—琥珀酰半醛还原酶 11—4-羟基丁酰辅酶A连接酶
12—4-羟基丁酰辅酶A脱氢酶 13—巴豆酰辅酶A水合酶 14—3-羟基丁酰辅酶A脱氢酶(NAD +)
15—乙酰乙酰辅酶A-酮硫解酶
目前,仅在古生菌勤奋金属球菌和Stygiolobus azoricus中确认了参与整个3-HP/4-HB循环过程中的所有酶,但该途径中的一些关键酶和关键基因在其他微生物也被检测到,如酸菌属(Acidianus spp.)和硫化叶菌属(Sulfolobus spp.)。此外,编码3-HP/4-HB途径特征酶的基因也存在于常温海洋古生菌群Ⅰ及Cenarchaeum symbiosum等微生物中,表明海洋中大量古生菌也存在利用该途径进行自养生长的可能。(https://www.daowen.com)
(6)二羧酸/4-羟基丁酸循环途径该途径(DB/4-HB)是由Huber等在研究嗜温性泉古生菌门除硫球菌目(Desulfurococcales)中的Ignicoccus hospitalis时发现并阐明的一条固定CO2的新途径,同时也存在于热变形菌目(Thermoproteales)古生菌。DB/4-HB途径也可以分为两个部分。第一部分是从乙酰辅酶A形成琥珀酰辅酶A阶段(图7-18左侧部分);第二部分是由琥珀酰辅酶A再生为乙酰辅酶A的过程(图7-18右侧部分)。本循环途径中涉及的主要代谢物有14种,包括乙酰辅酶A(乙酰-CoA)、丙酮酸、磷酸烯醇式丙酮酸、草酰乙酸、苹果酸、延胡索酸、琥珀酸、琥珀酰辅酶A、琥珀酰半醛、4-羟基丁酸、4-羟基丁酰辅酶A、巴豆酰辅酶A、3-羟基丁酰辅酶A及乙酰乙酰辅酶A。
图7-18 Desulfurococcales和Thermoproteales二羧酸/4-羟基丁酸循环途径
本途径中参与的酶:1—丙酮酸合成酶 2—丙酮酸/水二激酶 3—磷酸烯醇式丙酮酸羧化酶
4—苹果酸脱氢酶 5—延胡索酸水合酶 6—延胡索酸还原酶 7—琥珀酰辅酶A合成酶
8—琥珀酰辅酶A还原酶 9—琥珀酰半醛还原酶 10—4-羟基丁酰辅酶A连接酶
11—4-羟基丁酰辅酶A脱氢酶 12—巴豆酰辅酶A水合酶
13—3-羟基丁酰辅酶A脱氢酶(NAD +) 14—乙酰乙酰辅酶A-酮硫解酶
Fd—铁氧化还原蛋白
DB/4-HB途径第一部分中从草酰乙酸形成琥珀酰辅酶A的反应顺序及相关酶与逆向三羧酸循环中的相应反应顺序及酶一致(图7-15和图7-18),而从乙酰辅酶A到形成草酰乙酸的过程中所需的三种酶分别为丙酮酸合成酶、丙酮酸/水二激酶及磷酸烯醇式丙酮酸羧化酶。DB/4-HB途径第二部分反应顺序(从琥珀酰辅酶A到乙酰辅酶A再生)及所涉及的酶与3-HP/4-HB途径中的第二部分反应的顺序及相关酶一致。
2.微生物在有机物分解中的作用
有机物分解是指大分子有机物通过生物有氧呼吸彻底分解为CO2和H2O及通过无氧呼吸和发酵作用被降解为小分子有机物(如乙酸、乙醇及乳酸等)的过程。在无机物的有机化过程中,尽管一些自养微生物能利用多种途径将CO2固定并将其合成有机碳化物,但地球上有机碳化物的形成主要是依靠高等植物的光合作用;真菌和细菌等微生物是有机物分解的主力军。据报道,大气中90%以上的CO2主要来自于微生物对复杂有机碳化物及简单有机碳化物的分解,进而使得生物圈中的碳素得以平衡。
(1)简单有机物分解这里主要论述单糖及双糖的分解。在分解过程中,葡萄糖可以通过糖酵解(又称EMP)或其他途径形成丙酮酸,进而在有氧条件下经三羧酸循环(TCA)途径彻底氧化分解为CO2和H2O;也可以在无氧条件下经过发酵作用形成乙酸、乳酸、丁醇等小分子有机物。而其他单糖如果糖、半乳糖及甘露糖可以转变为糖酵解途径的中间代谢产物进入糖酵解途径。例如,果糖在果糖激酶及甘油醛激酶等酶的作用下生成甘油醛-3-磷酸,并进入糖酵解途径形成丙酮酸。同时,双糖在不同酶的作用下生成相应的单糖。例如,蔗糖在β-呋喃果糖苷酶的作用下生成果糖和葡萄糖,纤维二糖在纤维二糖磷酸化酶的作用下生成葡萄糖,麦芽糖在麦芽糖酶的作用下分解为葡萄糖,乳糖在β-半乳糖酶的作用下分解为半乳糖和葡萄糖(图7-19)。这些单糖在有氧条件下彻底氧化或在无氧条件下降解为小分子化合物。
丙酮酸的代谢去向因微生物种类及环境条件而异。微生物在有氧条件下丙酮酸可经过三羧酸循环彻底氧化为CO2和H2O;在无氧条件下,丙酮酸可经过不同微生物转化为多种发酵产物。不同微生物由丙酮酸出发的发酵途径可大致归纳为6条(图7-19)。
①酒精发酵:目前发现多种微生物可以发酵葡萄糖生成乙醇,包括酵母菌、霉菌及某些细菌。典型的酒精发酵是指酿酒酵母进行的同型酒精发酵,即葡萄糖先通过EMP途径降解为丙酮酸,丙酮酸在丙酮酸羧化酶作用下生成乙醛和CO2,乙醛在乙醇脱氢酶及其辅酶(NADH2)的作用下还原成乙醇的过程。此外,还有一些细菌也可以进行酒精发酵,例如,肠膜状明串珠菌(Leuconostoc mesenteroides)和发酵乳杆菌(Lactobacillus fermentum)可以利用HMP途径进行异型酒精发酵,生成乙醇、乳酸和CO2;运动发酵单孢菌(Zymomonas mobilis)及嗜糖假单胞菌(Pseudomonas saccharophila)利用ED途径先生成丙酮酸,然后进行同型酒精发酵生成乙醇。利用ED途径发酵生产乙醇的方法又称细菌酒精发酵。相比于传统的酵母酒精发酵,细菌酒精发酵具有代谢速率高、酒精得率高、代谢副产物少及不必定期供氧等优点。
②乳酸发酵:主要指一些革兰阳性、不产芽孢及微需氧的细菌利用糖类为底物且发酵终产物主要为乳酸的发酵。这类细菌称为乳酸菌。乳酸发酵根据发酵终产物种类可以分为同型乳酸发酵(homolactic fermentation)和异型乳酸发酵(heterolactic fermentation)。
③丙酸发酵:是指微生物利用葡萄糖、甘油或乳酸为底物进行发酵且产物主要为丙酸的发酵。用于丙酸发酵的菌种主要是丙酸杆菌属(Propionibacterium)。该属有多个种和亚种,均为厌氧菌。丙酸杆菌是一类革兰阳性、不产芽孢、不运动的细菌,菌落呈白色、黄色或者褐红色。一般情况下,最适pH为6.5~7.5,最适生长温度为28~37℃。国内外用于丙酸发酵研究的菌株主要集中在丙酸杆菌属的几个种,包括产酸丙酸杆菌(P.acidipropionici)、费氏丙酸杆菌(P.ffreudennreichii)、谢氏丙酸杆菌(P.shermanil)、特氏丙酸杆菌(P.thoenii)等。产酸丙酸杆菌利用葡萄糖、甘油和乳酸为底物时产生丙酸的总反应式分别为:
图7-19 微生物分解单糖及双糖途径
1.5C6H12O6→2CH3CH2COOH+CH3COOH+CO2+H2O
CH2OHCHOHCH2OH→CH3CH2COOH+H2O
3CH3CHOHCOOH→2CH3CH2COOH+CH3COOH+CO2+H2O
④混合酸发酵:进行此类发酵的微生物主要为肠杆菌科(Enterobacteriaceae)细菌,如埃希菌属(Escherichia)、沙门菌属(salmonella)和志贺菌属(Shigella)等。这些微生物经EMP途径将葡萄糖逐渐转化为多种有机酸,如甲酸、乙酸、乳酸及琥珀酸等,故称为混合酸发酵。发酵产物中除上述有机酸外,还有CO2、H2及2,3-丁二醇等。
⑤2,3-丁二醇发酵:自然界中具有产生2,3-丁二醇能力的微生物主要集中在克雷伯菌属(Klebisella)、芽孢杆菌属(Bacillus)及肠杆菌属(Enterobacter)中的一些种,如产酸克雷伯杆菌(Klebsiella oxytoca)、肺炎克雷伯杆菌(K.pneumoniae)、解淀粉芽孢杆菌(Bacillus amyloliquefaciens)、地衣芽孢杆菌(B.licheniformis)、多黏芽孢杆菌(B.polymyxa)及产气肠杆菌(Enterobacter aerogenes)等。其中产酸克雷伯杆菌和多黏芽孢杆菌显示出高产2,3-丁二醇的潜力;尤其是前者,同时具有宽广的底物谱及培养条件适应能力强的优点,因此产酸克雷伯杆菌常用于生物合成2,3-丁二醇。
⑥丁酸型发酵:能进行此类发酵的微生物主要为专性厌氧的梭状芽孢杆菌属(Clostridium),其中丁酸为其主要代谢产物。能产丁酸的梭状芽孢杆菌很多,其中高产丁酸的梭状芽孢杆菌有丁酸梭菌(Clostridium butyricum)和酪丁酸梭菌(C.tyrobutyricum)。此外,还有一些梭状芽孢杆菌的主要产物为丙酮及丁醇,其中目前研究最多的为丙酮丁醇梭菌(C.acetobutylicum)。梭状芽孢杆菌在生成上述主要产物的过程中,同时也会产生乙酸、乙醇、丙醇、CO2及H2等产物中的一种或几种。
(2)复杂有机碳化物的分解这里所指的复杂有机碳化物主要指含有碳及其他元素结构单元组成的高聚物分子。自然界中复杂有机碳化物的种类繁多,如纤维素、木质素、半纤维素、果胶质、淀粉等。对于复杂有机碳化物,微生物不能直接利用,而是通过种类繁多的微生物分泌相应的酶类使其分解为简单有机物,被微生物摄入胞内加以利用。分解纤维素、淀粉、半纤维素和木质素的微生物、水解酶类及分解过程详见本书第12章第1节内容。
果胶质是植物毗邻细胞之间的胞间层组分,由D-半乳糖醛酸通过α-1,4-糖苷键连接的直链构成,链上的羧基可部分或全部被甲醇氧化。根据是否含有甲基酯将果胶分为果胶酸和果胶酯。含甲基酯的果胶酯,可进一步与钙离子结合成不溶于水的原果胶。果胶酯酶有三类,即作用于果胶质甲基酯键并使果胶酯分解为果胶酸和甲醇的果胶质酯酶;水解α-1,4-糖苷键连接的直链结构的果胶质水解酶,其水解果胶酯的产物是D-甲基酯半乳糖单元及其寡聚体,水解果胶酸的产物则是D-半乳糖醛酸单元及其寡聚体;将果胶质裂解为变态的半乳糖醛酸单元的果胶质裂解酶。分解果胶质的真菌有青霉、曲霉、木霉、枝孢霉、根霉、毛霉等属的一些种。细菌中,好氧细菌以芽孢杆菌居多,如多黏芽孢杆菌(Bacillus polymyxa)、浸软芽孢杆菌(Bacillus carotovora);厌氧性细菌有蚀果胶梭菌(Clostridium pectinovorum)及费氏梭菌(Clostridium felsinneum)等。
(3)其他有机碳化物的分解脂肪是甘油和高级脂肪酸所形成的酯,不溶于水,可溶于有机溶剂。由饱和脂肪酸和甘油组成的、在常温下呈固态的称为脂;由不饱和脂肪酸和甘油组成的、在常温下呈液态的称为油。脂肪在微生物脂肪酶的作用下形成甘油和高级脂肪酸。甘油在甘油激酶及磷酸甘油脱氢酶的作用下可形成磷酸二羟丙酮,后者可进一步酵解成丙酮酸,其可以氧化为乙酰辅酶A,并进入三羧酸循环途径进一步彻底分解为CO2和H2O。脂肪酸则相对稳定,通常按照β-氧化的方式进行分解。在此过程中,高级脂肪酸被逐步分解为乙酸单体。在有氧条件下,乙酸被微生物氧化分解为CO2和H2O;在无氧条件下,乙酸则可被厌氧微生物转化为CH4和CO2。分解脂肪酸的好氧性细菌有假单胞菌、色杆菌(Chromobacterium)、无色杆菌(Achromobacter)及芽孢杆菌。真菌中有曲霉、芽枝霉和青霉。放线菌中的一些种也能分解脂肪。
自然界中存在多种烃类化合物,如植物叶面的蜡质、某些动植物表皮类脂质组分中的烃及石油中的烃类。微生物对烷烃分解的一般过程是逐步氧化,生成相应的醇、醛和酸,而后经过β-氧化进入三羧酸循环,最终分解成CO2和H2O。能够分解烃类的微生物有细菌、放线菌、霉菌、酵母菌等100余属,如分枝杆菌属、假单胞菌属、青霉属等。甲烷是最简单的烷烃。自然界能氧化甲烷的微生物主要是一群专性细菌,只能利用甲烷和甲醇作为碳源及能源而不能利用其他脂肪族碳氢化合物,如甲基孢囊菌属(Methylocystis)、甲基单胞菌属(Methylomomas)、甲基球菌属(Methylococcus)、甲基杆菌属(Methylobacterium)等不同属。
此外,环境中还存在着多种芳香化合物,如单宁、萜烯、生物碱、苯、甲苯、二甲苯、乙苯、萘酚、蒽及菲等。微生物能利用这些物质,但因底物结构与条件不同,分解程度与速度也各异。但其生物降解都具有一些共同的步骤,起始阶段都是改变或去除苯环上的可置换基团并导入羟基,常产生儿茶酚、原儿茶酚和龙胆酸等共同中间产物,这些产物被开环分解,最终彻底氧化成二氧化碳和水。参与分解芳香化合物的细菌主要有假单胞菌属(Pseudomonas)、分枝杆菌属(Mycobacterium)、不动杆菌属(Acinetobacter)等属;真菌中的杂色云芝(Coriolus versicolor)、青霉属(Penicillium)、曲霉属(Aspergillus)等都能分解芳香族化合物。
二、氮循环
氮元素是核酸和蛋白质的主要成分,是构成生物体的必要元素。自然界氮元素含量丰富,主要以分子态氮(N2)、有机氮化合物(R-NH2)、氨(NH3)和铵盐( )、亚硝酸盐(
)、亚硝酸盐( )、硝酸盐(
)、硝酸盐(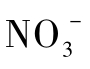 )5种形式存在。尽管分子态氮在大气中的含量很丰富(约79%,体积分数),但所有动植物及大多数微生物无法直接利用分子态氮。有机态氮必须通过微生物的分解才能被绿色植物等初级生产者利用。而自然界中能被植物吸收利用的高度水溶性的无机氮化合物(铵盐、硝酸盐等)数量却很少,远远不能满足地面植物对氮素营养的需要。因此,只有将不同形态的氮互相转化和循环,才能满足所有生物体对氮素营养的需求。
)5种形式存在。尽管分子态氮在大气中的含量很丰富(约79%,体积分数),但所有动植物及大多数微生物无法直接利用分子态氮。有机态氮必须通过微生物的分解才能被绿色植物等初级生产者利用。而自然界中能被植物吸收利用的高度水溶性的无机氮化合物(铵盐、硝酸盐等)数量却很少,远远不能满足地面植物对氮素营养的需要。因此,只有将不同形态的氮互相转化和循环,才能满足所有生物体对氮素营养的需求。
氮素循环包括固氮作用、氨化作用、硝化作用及反硝化作用等转化作用(图7-20)。大气中的分子态氮被固氮微生物(如根瘤菌、蓝细菌、巴氏固氮梭菌等)转化为氨,一部分氨可以直接被微生物和植物同化为有机氮化合物,一部分还可以被硝化细菌氧化成硝酸盐,后者通过植物吸收利用将无机氮化合物转化为有机氮化合物。存在于微生物及植物体的含氮化合物被动物食用后转化为动物蛋白。最后,动植物和微生物的残体及其排泄物中的有机氮化物被微生物分解时转化为氨,或被氧化成硝酸盐供植物吸收利用,或被进一步转化为气态氮返回大气中,这样循环往复构成了氮素在生态系统中的完整循环。此外,还有非生物过程使氮进入生态系统。火山活动释放出沉积矿床中的氮,自然电离(如闪电)、机动车排气释放出氮氧化合物到空气中;大气中一些含氮物最终通过沉积作用进入生态系统。
图7-20 生态系统中氮元素循环
1.固氮作用
固氮作用(nitrogen fixation)是指将大气中的分子态氮转化为氨,进而合成有机氮化物的过程,分为生物固氮和非生物固氮两种方式。生物固氮(约占到全球固氮量的90%)是自然界氮循环的重要环节,相当于工业固氮量的2.5倍。在现代农业中,生物固氮不仅具有增强土壤肥力及提高农作物产量的作用,而且对维持整个自然界生态平衡具有重要意义。生物固氮主要是指微生物固氮。目前已报道的固氮微生物达100多属,近200种。它们都是原核微生物,包括细菌、蓝细菌、放线菌和古生菌等。根据固氮微生物的生态类型可以分为自生固氮菌、共生固氮菌和联合固氮菌三类。
自生固氮菌是指一类能自由地生长在海洋与土壤等环境中且能独立进行固氮的微生物。根据营养方式自生固氮菌又可分为化能自养、光能自养、化能异养和光能异养固氮菌四类。化能自养固氮菌如产碱菌属(Alcaligenes)和硫杆菌属(Thiobacillus)中的氧化亚铁硫杆菌(T.ferrooxidans)。光能自养固氮菌可分为非产氧的光合细菌和产氧的蓝细菌两大类群。光合细菌类群广泛存在于海域和淡水环境,如着色菌属(Chromatium)和绿假单胞菌属(Chloropseudomonas)。蓝细菌在自然界中分布广泛,许多种属能进行固氮作用,固氮蓝细菌固氮量仅次于豆科植物与根瘤菌的共生固氮系统。现已证明24个属的120多个种具有固氮能力,其中大多属于念珠藻科(Nostocaceae),少数属于胶须藻科(Rivulaiaceae)等。化能异养固氮菌有克雷伯菌属(Klebsiella)、固氮菌属(Azotobacter)、拜叶林克菌属(Beijerinckia)、芽孢梭菌属(Clostridium)、甲烷八叠球菌属(Methanosarcina)及芽孢杆菌属(Bacilllus)中的一些菌种。光能异养固氮菌如光合细菌中的红螺菌属(Rhodospirillum)和红假单胞菌属(Rhodopseudomonas)中的一些菌种。
共生固氮菌是指必须与它种生物共生在一起才能固氮的微生物。这类固氮微生物包括与豆科植物共生的根瘤菌,如根瘤菌属(Rhizobium)、固氮根瘤菌属(Azorhizobium)、中华根瘤菌属(Sinorhizobium)等;与非豆科植物共生的弗兰克菌属(Frankia);与水面蕨类植物红萍共生的红萍鱼腥藻(Anabaena azollae)。地衣是蓝细菌和真菌的共生体,具有固氮作用的蓝细菌有念珠蓝细菌属(Nostoc)及鱼腥蓝细菌属(Anabaena)等。
联合固氮是介于典型的自生固氮和共生固氮之间的一种中间类型,又称“弱共生”或“半共生”固氮作用。相对于普通自生固氮作用,联合固氮的专一性强且固氮作用强;但与典型共生固氮的不同是不形成根瘤、叶瘤那样独特的形态结构。这类固氮微生物必须生活在植物根际、叶面或动物肠道等环境中才能进行固氮,如生脂固氮螺菌(Azospirillum lipoferum)和肠杆菌属(Enterobacter)。此外,许多自生固氮菌也可以进行联合固氮,如克雷伯菌属、拜叶林克菌属、固氮菌属和芽孢杆菌属等。
各类固氮微生物进行固氮的基本反应式相同,固氮过程为与固氮酶结合的分子态氮被逐级还原的过程,最终生成氨(图7-21)。固氮过程必须满足以下基本条件:①必须有具固氮活性的固氮酶;②必须有电子和质子供体,每还原1分子N2需要6个电子和6个质子,另有2个质子和电子用于生成H2,为了传递电子和质子,还需要相应的电子传递链;③必须有能将N2分子三键打开的能量;④有严格的无氧环境或保护固氮酶的免氧失活机制;⑤形成的氨必须及时转运或转化排出,避免氨反馈阻抑效应。
图7-21 生物固氮的总反应式(上)及固氮原理示意(下)
2.氨化作用
微生物分解有机氮化合物产生氨的过程称为氨化作用(ammonification)。有机氮化合物的种类较多,主要是蛋白质及水解后的氨基酸、尿素、几丁质和尿酸等。具备较强氨化能力的微生物称为氨化菌,主要有芽孢杆菌、变形杆菌、假单胞菌、芽孢梭菌、放线菌及曲霉、青霉、根霉、毛霉等。氨化作用产生的氨一部分经微生物和植物同化作用合成为有机氮,一部分经硝化作用逐步转化为硝酸盐。蛋白质在胞外被蛋白酶水解成小分子肽、氨基酸后才能进入细胞被吸收利用。微生物分解氨基酸的主要方式为脱羧作用和脱氨作用,其中氨基酸脱氨作用的方式分为氧化脱氨、还原脱氨、水解脱氨及去饱和脱氨。核酸的氨化作用是指核酸在胞外核糖核酸酶、胞外脱氧核糖核酸酶等酶的作用下分解为嘌呤或嘧啶,后者被多种微生物(如诺卡菌、假单胞菌、小球菌和梭状芽孢杆菌等)进一步分解并最终形成氨基酸、尿素及氨的过程。几丁质是一种含氮多聚糖,其基本结构单位为N-乙酰葡萄糖胺,广泛存在于真菌细胞壁,尤其是很多担子菌。几丁质分解是通过微生物分泌的几丁质酶及几丁二糖酶将长链切割成几个单位的短链寡糖胺、几丁二糖、N-乙酰葡萄糖胺、葡萄糖胺及乙酸等。葡萄糖胺最后脱氨基成为葡萄糖和氨。能够分解几丁质的微生物以放线菌为主,如链霉菌属、诺卡菌属、小单胞菌属、游动放线菌属和胞囊链霉菌属等。分解尿素的微生物种类繁多,广泛分布于土壤和污水池,尤其是在粪尿池及堆粪场上。常见的分解尿素的细菌包括芽孢杆菌、小球菌、假单胞菌、棒状杆菌、梭状芽孢杆菌及尿细菌(如Urobacillus pasteurii)。这些微生物能将尿素分解为氨。人和动物尿中的尿酸和马尿酸,在微生物作用下可被分解,生成的氨基酸按上述脱氨方式转化为氨。马尿酸的分解反应式见图7-22。
图7-22 马尿酸的分解反应式
3.硝化作用
硝化作用(nitrification)是指微生物在有氧条件下将氨态氮氧化成硝态氮的过程。整个硝化作用过程分为氨态氮被氧化成亚硝酸盐及亚硝酸盐被氧化为硝酸盐两步反应,其中NH2OH为氨氧化过程的第一个中间产物(图7-23)。
参与硝化作用第一步反应的细菌称为亚硝酸细菌或铵氧化菌(ammonia oxidizer),如亚硝化单胞菌属(Nitrosomonas)、亚硝化螺菌属(Nitrosospira)、亚硝化球菌属(Nitrosococcus)等革兰阴性菌。亚硝化单胞菌普遍存在于土壤中,为最早分离的亚硝酸细菌,标准种为欧洲亚硝化单胞菌(N.europaea)。亚硝化单胞菌细胞杆状,多以单个存在,很少呈链状,大小为(0.8~0.9)μm×(1.0~2.0)μm,具有两根极生鞭毛,严格化能无机营养,能将铵和羟胺氧化成亚硝酸,最适生长温度为25~30℃,最适pH为7.5~8.0。参与硝化作用第二步反应的细菌称为硝酸细菌或亚硝酸氧化菌(nitrite oxidizer)。最早定名的硝酸细菌为硝化杆菌属(Nitrobacter),标准菌株为维氏硝化杆菌(N.winogradskyi),细胞杆状,在液体中生长时细胞单个或成堆,外围有黏质,呈楔形或梨形,大小为(0.6~0.8)μm×(0.6~0.8)μm,通常不运动。自然界中除了化能自养菌为硝化作用的主要推动者外,还有化能异养菌参与硝化作用,如细菌及真菌中的曲霉等部分化能异养菌也具有将氨氧化成亚硝酸和硝酸的能力。
图7-23 硝化作用反应过程及相关酶
AMO—氨单加氧酶HAO—羟胺氧化还原酶NXR—亚硝酸氧化还原酶UQ—醌 UQH2—全醌
4.硝酸盐还原与反硝化作用
硝酸盐还原方式可以分为同化硝酸盐还原和异化硝酸盐还原两种。同化硝酸盐还原是指硝酸盐被还原为氨和亚硝酸盐,氨又被同化成氨基酸的过程;异化硝酸盐还原是指微生物在无氧或微氧条件下进行的硝酸盐呼吸,即以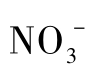 或
或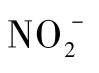 代替O 2作为电子受体进行的呼吸代谢。异化硝酸盐还原又分为发酵型硝酸盐还原(fermentative nitrate reduction)和呼吸型硝酸盐还原(respiratory nitrate reduction)。呼吸型硝酸盐还原又称为反硝化作用,即微生物将硝酸盐(
代替O 2作为电子受体进行的呼吸代谢。异化硝酸盐还原又分为发酵型硝酸盐还原(fermentative nitrate reduction)和呼吸型硝酸盐还原(respiratory nitrate reduction)。呼吸型硝酸盐还原又称为反硝化作用,即微生物将硝酸盐(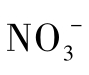 )依次还原为亚硝酸盐(
)依次还原为亚硝酸盐( )、一氧化氮(NO)、一氧化二氮(N2O)及氮气(N2)的过程。在反硝化过程中,硝酸盐经一系列酶的作用,以细胞色素传递电子,最终被还原为N2O及氮气(N2)并释放到大气中。该过程是一个偶联产能的过程,一分子硝酸盐被还原后能产生2分子的ATP。能进行硝酸盐呼吸的都是一些兼性厌氧菌即反硝化细菌,如假单胞菌属(Pseudomonas)、芽孢杆菌属(Bacillus)、硫杆菌属(Thiobacillus)及丙酸杆菌属(Propionibacterium)等。反硝化过程涉及多种酶类,主要包括硝酸还原酶、亚硝酸还原酶、一氧化氮还原酶及一氧化二氮还原酶。多数反硝化细菌具有还原硝酸至氮气所必需的全部还原酶类,终端产物为N2O及N2。但也有一些反硝化细菌缺少还原酶,只能以
)、一氧化氮(NO)、一氧化二氮(N2O)及氮气(N2)的过程。在反硝化过程中,硝酸盐经一系列酶的作用,以细胞色素传递电子,最终被还原为N2O及氮气(N2)并释放到大气中。该过程是一个偶联产能的过程,一分子硝酸盐被还原后能产生2分子的ATP。能进行硝酸盐呼吸的都是一些兼性厌氧菌即反硝化细菌,如假单胞菌属(Pseudomonas)、芽孢杆菌属(Bacillus)、硫杆菌属(Thiobacillus)及丙酸杆菌属(Propionibacterium)等。反硝化过程涉及多种酶类,主要包括硝酸还原酶、亚硝酸还原酶、一氧化氮还原酶及一氧化二氮还原酶。多数反硝化细菌具有还原硝酸至氮气所必需的全部还原酶类,终端产物为N2O及N2。但也有一些反硝化细菌缺少还原酶,只能以 为电子受体;而另一些又缺乏N2O还原酶,导致N2O为其终端产物;还有一些反硝化细菌虽具有N2O还原酶,但不能以
为电子受体;而另一些又缺乏N2O还原酶,导致N2O为其终端产物;还有一些反硝化细菌虽具有N2O还原酶,但不能以 和
和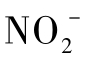 为电子受体产生N2O。
为电子受体产生N2O。
反硝化作用会引起土壤氮肥的严重损失,甚至达到施入化肥量的75%左右。影响反硝化作用的因素很多,主要有土壤中的水分、pH、温度、通气量、有机碳含量、耕作及硝态氮的浓度和施氮量等。一般来讲,反硝化速率与pH呈正相关(最适宜pH为7.0~8.0),但与氧分压呈负相关。
三、硫循环
硫是一些必需氨基酸、维生素和辅酶的必需元素,是生命有机体的重要组成部分,约占干重的1%。自然界中的硫以单质硫、无机硫化物和有机态硫的形式存在,其中有机态硫占50%~75%。生态系统中各种形态的硫可在化学和生物的作用下相互转化(图7-24),构成了硫循环,且每个环节都有相应的微生物参与。
图7-24 生态系统中硫元素循环
1.有机态硫分解
有机态硫分解是指动植物和微生物残体中的有机态硫(如蛋白质),被微生物降解为无机硫化物的过程,又称脱硫作用。一般来讲,凡能将含氮有机物分解产氨的氨化微生物都具有脱硫作用,相应的氨化微生物也可称为脱硫微生物。如变形杆菌将半胱氨酸水解为氨和硫化氢。在有氧条件下,微生物将有机态硫转化为H2S,并逐步氧化为硫酸盐,进而为植物和微生物提供硫营养。在无氧条件下,特别是蛋白质物质腐烂分解时,积累H2S和有气味的硫醇如硫甲醇(CH3SH)于环境中,一旦超过某种浓度可危害植物和其他生物。土壤中积累H2S较多时,对植物根部有毒害作用。
2.无机硫同化作用
微生物利用外源和H2S合成自身增殖及代谢所需含硫细胞物质的过程称为硫的同化作用。大多数细菌和真菌都能像植物一样利用硫酸盐作为唯一硫源,转变为巯基的形式固定在蛋白质等有机物中。上述过程又称为同化硫酸盐还原作用,即硫酸盐中的硫由正六价氧化态转变为负二价还原态。只有少数微生物能同化H2S,大多数情况下单质硫和H2S等都需要先转变为硫酸盐,再固定为有机态硫。
3.硫氧化作用
硫氧化作用又称硫化作用,是指还原态的无机硫被微生物氧化为硫酸或其中的部分氧化化合物的过程,如微生物可将S氧化为硫代硫酸盐,或 被氧化为
被氧化为 (图7-25)。参与硫氧化作用的微生物主要有硫化细菌及硫磺细菌。硫化细菌属于革兰阴性的硫杆菌属(Thiobacillus),如氧化硫硫杆菌(T.thiooxidans)、排硫硫杆菌(T.thioparus)、氧化亚铁硫杆菌(T.ferrooxidans)和脱氮硫杆菌(T.denitrificans)等。该类群微生物能氧化H2S、单质S、
(图7-25)。参与硫氧化作用的微生物主要有硫化细菌及硫磺细菌。硫化细菌属于革兰阴性的硫杆菌属(Thiobacillus),如氧化硫硫杆菌(T.thiooxidans)、排硫硫杆菌(T.thioparus)、氧化亚铁硫杆菌(T.ferrooxidans)和脱氮硫杆菌(T.denitrificans)等。该类群微生物能氧化H2S、单质S、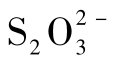 及
及 等无机硫化物,并产生能量、硫酸及同化CO2合成有机物。硫磺细菌是一类能将H2S氧化为S并将硫积累在细胞内的细菌,包括丝状硫细菌和光能自养硫细菌。丝状硫细菌有贝日阿托菌属(Beggiatoa)、透明颤菌属(Vitreoscilla)、辫硫菌属(Thioploca)及亮发菌属(Leucothrix)等。光能自养硫细菌主要为一些有色硫细菌,如紫硫菌和绿硫菌。
等无机硫化物,并产生能量、硫酸及同化CO2合成有机物。硫磺细菌是一类能将H2S氧化为S并将硫积累在细胞内的细菌,包括丝状硫细菌和光能自养硫细菌。丝状硫细菌有贝日阿托菌属(Beggiatoa)、透明颤菌属(Vitreoscilla)、辫硫菌属(Thioploca)及亮发菌属(Leucothrix)等。光能自养硫细菌主要为一些有色硫细菌,如紫硫菌和绿硫菌。
图7-25 单质硫氧化(1)及亚硫酸盐氧化(2)
Sn—聚合态硫 GSH—还原型谷胱甘肽 GSSG—氧化型谷胱甘肽 CytC—细胞色素C
4.反硫化作用
反硫化作用是指当环境处于缺氧状态时,硫酸盐、亚硫酸盐、硫代硫酸盐等高价态硫在微生物作用下被还原为硫化氢的过程。其中硫被微生物还原成硫化氢的过程称为异化性硫还原作用;硫酸作为厌氧菌电子传递链的末端电子受体而被还原为亚硫酸或硫化氢的过程称为异化性硫酸盐还原作用,脱硫菌属(Desulfobacter)及脱硫弧菌属(Desulfovibrio)细菌能进行该反应。脱硫脱硫弧菌(Desulfovibrio desulfuricans)利用葡萄糖和乳酸还原硫酸盐的过程见图7-26。
图7-26 脱硫脱硫弧菌利用葡萄糖和乳酸还原硫酸盐的过程
四、磷循环
磷是一切生命体的重要营养元素,是生物膜、遗传信息载体(DNA)、高能量化合物及体内糖代谢的某种中间化合物的重要组成部分。磷主要以三种状态存在,水溶性无机磷(如磷酸二氢盐)、有机磷(如核酸、植酸及卵磷脂等)和存在于沉积物中的不溶性或微溶性的无机磷。上述三种状态的磷在生物圈中不断相互转化,进而构成了维持磷元素平衡的磷循环(图7-27)。该循环可分为不溶性无机磷的可溶化、可溶性无机磷的有机化及有机磷矿化三个主要环节。不溶性无机磷的可溶化主要是指土壤和岩石中不溶性含磷化合物(如钙、镁磷酸盐)在微生物(如硝化细菌和硫化细菌)所产有机酸或无机酸的作用下形成可溶性含磷化合物的过程。可溶性无机磷的有机化是指可溶性含磷化合物经动物、植物和微生物吸收被同化为有机磷的过程。有机磷矿化是指生物体中的有机磷化合物被相应的微生物分解为可供生物重新利用的可溶性无机磷化合物的过程。参与有机磷矿化的微生物包括细菌、放线菌及真菌,如芽孢杆菌属、链霉菌属、假单胞菌属、曲霉和青霉属的一些微生物。
图7-27 生物圈中磷元素循环
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。



