
1.SP-C分子结构及其在肺泡磷脂单分子膜中的行为特性
由综述已知,SP-C是由35个氨基酸组成的高度疏水性蛋白,并且它是唯一的完全由肺泡Ⅱ型上皮细胞分泌的且仅存在于肺泡中的保守性结合蛋白。虽然SP-C在肺泡中的含量较少,但它是维持肺泡正常功能的必不可少的部分。在肺泡内,SP-C能够与磷脂相互作用,在呼吸压缩过程中促使磷脂分子从单分子膜上挤出,在舒展过程中又促进底液中的磷脂重新吸附到单分子膜上。根据SP-C的分子结构,它在肺泡单分子膜中的行为特性如下。
(1)SP-C的Cys-5,Cys-6氨基酸残基通过硫酯键与两条棕榈酰链结合,由于这两条棕榈酰链为疏水链,可插入肺泡磷脂单分子膜内,从而增加SP-C分子在单分子膜中的稳定性。
(2)SP-C的Lys-11,Arg-12为碱性氨基酸,在生理pH值下,其电离带正电,更易与带负电的磷脂PG结合。实验表明,SP-C对带负电的PG脂膜的影响大于对PC脂膜的影响。因此,单分子膜中的SP-C分子周围的PG成分较多。对于PG中的DPPG磷脂来说,其极性头基间存在的负电排斥力易形成LE相膜,但由于其为饱和磷脂,所以高表面张力下也不易挤出。然而实验表明,SP-C的存在会促进DPPG的挤出。
(3)SP-C的第9~34氨基酸残基序列由富含氨酸(Val)的疏水氨基酸组成,它们形成α-螺旋结构,长度约为37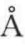 ,稳定且具有刚性。SP-C独特的α-螺旋结构与两条棕榈酰链赋予了SP-C的极度疏水性。在肺泡中,它或与磷脂分子结合存在于胞液中,或吸附到单分子膜表面,其中α-螺旋结构在单分子膜上与界面的倾角为20。左右。
,稳定且具有刚性。SP-C独特的α-螺旋结构与两条棕榈酰链赋予了SP-C的极度疏水性。在肺泡中,它或与磷脂分子结合存在于胞液中,或吸附到单分子膜表面,其中α-螺旋结构在单分子膜上与界面的倾角为20。左右。
(4)在模拟肺泡单分子膜的实验中,FLM技术显示单分子膜在压缩初期为LC相(liquid condensed state)与LE相(liquid expanded state)两相共存,继续压缩单分子膜时LE相成分(主要为非DPPC成分)发生挤出。Amrein等通过SFM技术分析表明,被挤出成分没有分散到底液中,而是通过SP-C分子与单分子膜紧密相连;当单分子膜舒展时,挤出成分又重新插入单分子膜。Edwin Veldhuizen认为,SP-C的挤出是指α-螺旋结构的挤出,而SP-C的Cys-5,Cys-6位结合的两条疏水性棕榈酰链不随α-螺旋结构的挤出而挤出。
2.SP-C/磷脂的行为模型
综上所述,肺泡单分子膜在压缩舒展过程中,SP-C与磷脂成分的挤出及再插入密切相关。根据以上SP-C分子的行为特性,我们提出以下四点假设。
(1)肺泡单分子膜中SP-C周围富集PG成分,单分子膜压缩时,不饱和PG与SP-C的α-螺旋结构自发地发生挤出。此过程引起能量的转变,从而促进DPPG成分挤出。
(2)在单分子膜压缩过程中,当LE相达到崩溃表面压时,不饱和PG首先形成崩溃凹槽,随后SP-C的α-螺旋结构受凹点区的吸引,随磷脂分子一同迅速被挤出,在底液中形成磷脂-蛋白质聚集体。此聚集体是以α-螺旋结构为中心,脂类分子聚集在四周的微囊结构。SP-C通过两条疏水棕榈酰链与表面单分子膜保持连接。
(3)在单分子膜舒展过程中,膜表面积增加,表面张力增大,受表面张力的影响,上述磷脂-蛋白质聚集体结构被打破,并迅速吸附到单分子膜表面。(www.daowen.com)
(4)在挤出及插入过程中,α-螺旋结构的角度发生变化,SP-C的Pro-7氨基酸残基构象存在正反式构象间的变换。
根据以上假设,提出以下SP-C/磷脂相互作用的行为模型(见图4-19)。
由图4-19可知:SP-C与磷脂在气-液表面形成单分子膜。DPPC与PG由于部分互溶,局部分布不均,单分子膜由扩展膜(图4-19中的Ⅰ)压缩时,发生相分离。DPPC富集区为LC相,PG富集区为LE相(图4-19中的Ⅱ)。膜继续压缩,当LE相达到崩溃表面压时,不饱和磷脂首先形成崩溃凹槽(图4-19中的Ⅲ),而后SP-C的α-螺旋结构受凹槽区的吸引,随磷脂一同迅速被挤出,在底液中形成以SP-C的α-螺旋结构为中心的磷脂-蛋白质集合体(图4-19中的Ⅳ)。由于LE相区非DPPC成分的挤出,单分子膜表面DPPC成分得到富集,此后再继续压缩膜,表面张力可降至最低。Schürch等利用电子显微镜,在小鼠肺中观测到由肺表面活性物质组成的多层结构。这些结构可能会在压缩过程中储存表面活性物质,因此被称作表面结合储存体。M.Amnrein等认为这个储存体是一个富集非DPPC成分的脂双层结构。在这个脂双层结构中,SP-C与磷脂分子间的相互作用关系可能和生物膜脂双层中蛋白质与磷脂的关系相类似,即著名的“流动镶嵌模型”。这种脂双层结构是相当稳定的,因此在舒展过程中,脂双层结构不易被打破,而其中的分子也不能迅速吸附到单分子膜表面。与脂双层结构相比较,我们提出的磷脂-蛋白质聚集体结构更加合理,它是一个亚稳态结构,具有更高的能量和更大的流动性。在单分子膜舒展过程中,随着表面张力的增大,磷脂-蛋白质聚集体结构易被打破(图4-19中的Ⅴ),其中的分子可以迅速地重新插回表面的单分子膜中(图4-19中的ⅥD。SP-C及周围的磷脂重新插入单分子膜上,从而使刚性脂DPPC流化,加速单分子膜的铺展,单分子膜重新回到初始状态(图4-19中的Ⅰ)。
图4-19 SP-C/磷脂在肺泡单分子膜中相互作用的行为模型
由假设模型可知,SP-C在肺泡压缩舒展循环过程中,起到以下关键性的作用。
(1)在单分子膜压缩时,SP-C不断蓄积能量并促进非DPPC成分挤出,使单分子膜表面张力可达0。
(2)挤出的磷脂以SP-C为中心,形成磷脂储存区,并通过SP-C的棕榈酰链与单分子膜连接。
(3)在单分子膜扩张过程中,SP-C释放能量,促进磷脂储存区成分及时插回膜表面,加速DPPC的铺展,减少吸气阻力。
因此,SP-C的存在调节了肺泡单分子膜中磷脂成分的含量,起到能量枢纽的作用,使表面单分子膜在呼吸过程中保持稳定。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。





